Классификация
летнего населения птиц равнин Северной Евразии
опубликована: Сибирский
экологический журнал, 2001, т. 8, No 6, с. 741-766.
Авторы: Е.С.Равкин1),
Ю.С.Равкин2), Л.Г.Вартапетов2), И.Н.Богомолова2),
К.В.Торопов2), С.М.Цыбулин2), В.С.Жуков2), И.В.Покровская3),
И.И.Бышнев4), С.К.Кочанов5)
1) МосНПО "Радон"
127562 Москва,
Алтуфьевское шоссе, 30-229
2) ИСиЭЖ СО РАН
630091 Новосибирск, ул.
Фрунзе, 11
3) ИГ РАН
109017 Москва,
Старомонетный пер., 29
4) Березинский заповедник
211188 Беларусь, Витебская
область, Лепельский район, Домжерицы
5) ИБ КНЦ УрО РАН
Сыктывкар,
ул.Коммунистическая, 28
АННОТАЦИЯ
Сопоставление
результатов кластерного анализа летнего населения птиц, проведенного отдельно и
в совокупности по Восточно-Европейской и Западно-Сибирской равнинам, показало
значительные различия в объемах и границах распространения концептуальных типов
орнитокомплексов. При этом границы типов и других таксонов классификаций не
совпадают с ландшафтными и геоботаническими рубежами, хотя во всех случаях
определяются сходным набором факторов среды. По обобщенной классификации в
природных и полевых сообществах птиц суши южная граница северной надтиповой
группировки (между лесотундровым и лесным типом населения) в Западной Сибири
проходит между северной и средней тайгой, а в Европе - в пределах северной
тайги. В Европе, в результате отепляющего влияния Северо-Атлантического
течения, она вслед за растительностью смещена к северу, а в Сибири из-за
значительной площади болот и пойм, а также из-за разреженности северотаежных
лесов гипоарктические виды птиц и, соответственно, рубеж между лесотундровым и
лесным типами населения существенно смещены к югу, вплоть до границы между
северной и средней тайгой. Северная граница лесотундрового населения в западной
части Восточной Европы перемещается почти к берегам Северного Ледовитого океана
в соответствии с диагональным смещением типа растительности по направлению к
северу.
ВВЕДЕНИЕ
Население птиц (орнитокомплексы, сообщества) в рамках парадигмы факторной
зоогеографии до недавних пор описывали и анализировали по природным участкам,
провинциям, подзонам или зонам [1-10 и др.]. С накоплением материалов появились
обобщения на уровне физико-географических стран [11-14], и в самое последнее
время выполнены сравнения по Восточно-Европейской и Западно-Сибирской равнинам
[15, 16]. В предлагаемой работе впервые приводится единая (обобщенная)
классификация орнитокомплексов, выполненная на основе кластерного анализа
материалов по обеим равнинам. При этом ранее прирост объема анализируемого
материала приводил к достраиванию классификационных представлений. В данном
случае при объединении произошли существенные изменения в представлениях о
неоднородности орнитокомплексов. Такие отличия при использовании кластерного и
факторного анализов описаны ранее. Относительно населения птиц они
свидетельствуют о не устоявшихся представлениях об орнитологической специфике
понятия типа сообществ, рабочем характере выполненных классификаций, их
регионально-провинциальной (зонально-подзональной) ограниченности.
МАТЕРИАЛЫ И МЕТОДЫ
Для составления обобщенной классификации населения птиц Восточно-Европейской и
Западно-Сибирской равнин использованы авторские материалы, накопленные в банке
данных лаборатории зоологического мониторинга ИСиЭЖ СО РАН. Кроме того, взяты
сведения из литературных источников (около 70) и данные других вкладчиков
банка, любезно предоставивших их для рассмотрения. Библиография публикаций по
использованным материалам не приведена из-за ограниченности объема статьи. Все
участники и районы работ перечислены в наших прежних публикациях [11, 13, 15,
17]. Учеты птиц проведены в первой половине лета (с 16 мая по 15 июля) в
1880-1907, 1936-1938, 1947-1997 гг., главным образом с конца 1970-х гг.
Исследованиями охвачены основные по площади местообитания во всех природных
зонах Восточно-Европейской равнины, а также российской части Фенноскандии и
Западно-Сибирской равнины (в пределах территории СССР на 1990 г.). Следует
отметить, что Восточно-Европейская равнина по принятому нами физико-географическому
районированию [17] не включает полупустыни и пустыни Прикаспийской низменности.
Всего для расчетов использовано 2649 вариантов орнитокомплексов. Под вариантом
населения под-разумевается комплекс всех видов птиц, встреченных в данном
местообитании за один сезон с учетом их обилия (особей/км2).
Несмотря на значительные объемы
собранных материалов, на результатах анализа иногда сказывается неравномерность
охвата территории. Так, в Европе тундровая и степная зоны обследованы хуже, чем
лесная и лесостепная. По водно-околоводным и рудеральным сообществам меньше
данных и по Европе, и по Сибири. Отчасти поэтому видовое обилие по вариантам
населения усреднено по наименьшим выделам карт растительности европейской части
СССР, Кавказа (1987) и Западно-Сибирской равнины (1976) масштабов 1 : 2 000 000
и 1 : 1 500 000. В соответствии с этим материалы объединены в 536 групп. Такое
усреднение в некоторой степени нивелирует неравномерность в объемах собранных
сведений, методические различия, а также годовые и индивидуальные отличия
вариантов населения.
По рассчитанным средним вычислены
коэффициенты сходства Жаккара [18] для количественных признаков [19]. Эти
коэффициенты использованы для факторной классификации с поэтапной доразбивкой
крупных классов, состоящих из пяти и более средних. Классификация проведена с
помощью одного из методов кластерного анализа [20]. При этом для составления
концептуальной (итоговой) классификации использованы только интерпретируемые и
понятные с предметной точки зрения разбиения [16, 21]. Поэтому дробность
деления разных таксонов неодинакова. Подробнее методологические и методические
основы исследований опубликованы ранее [4, 11]. Названия птиц даны по А. И.
Иванову [22], типы фауны - по Б. К. Штегману [23] с некоторыми изменениями и
дополнениями.
Классификации населения птиц
отдельно по Западно-Сибирской и Восточно-Европейской равнинам опубликованы
ранее [13, 16]. Следует учитывать, что обобщенные представления по всей
территории получены не объединением двух классификаций, а по расчетам,
выполненным заново по объединенным материалам. Поэтому границы в едином
варианте не повторяют таковые в отдельных классификациях, а проведены там, где
это целесообразно, исходя из разделения всей совокупности данных.
НЕКОТОРЫЕ ОСОБЕННОСТИ
КЛАССИФИКАЦИИ И ТИПОЛОГИИ ЖИВОТНОГО НАСЕЛЕНИЯ
Поскольку понятие типа животного населения определено недостаточно четко, а
объем его варьирует, имеет смысл кратко сформулировать некоторые представления
о типологии и классификации в целом и типе в частности, которые подробнее
опубликованы ранее [24].
Известно, что результат
формализованной классификации зависит от представительности имеющегося
материала. При разной полноте данных классификация может приводить к различным
результатам. Так, на рисунке показано, что если анализируются пробы с 1-й по
4-ю, то 2-я и 3-я ближе друг к другу, чем к 1-й и 4-й, и будут относиться по
сходству к одному таксону. Первая и четвертая пробы менее сходны и образуют
другие таксоны. При бoльшем объеме (1-6) пробы 2 и 3 последовательно ближе к пробам
5 и 6 и должны быть отнесены к разным таксонам. При еще бoльшем объеме данных
(1-7) они снова разойдутся в разные таксоны, поскольку сходство проб 1-4 больше
между собой, чем с пробами 5-7. Последние, в свою очередь, ближе друг к другу,
и основная разделительная линия идет ниже их, отличая друг от друга эти
подсовокупности.
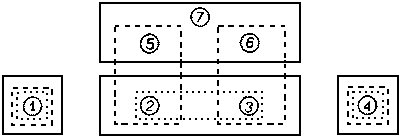
Зависимость результата классификации от представительности имеющихся
материалов.
Совокупности проб: точки - 1-4, тире - 1-6, непрерывной линией - 1-7.