Численность, распределение и
пространственно-типологическая неоднородность населения земноводных и
пресмыкающихся в Томской и Новосибирской областях
Ю.С.Равкин, В.Н.Куранова,
С.М.Цыбулин, И.Н.Богомолова, В.А.Юдкин,
К.В.Торопов, Н.А.Булахова, О.Б.Борисович, В.В.Панов,
А.К.Добротворский
Институт систематики и экологии
животных СО РАН, Новосибирск
Томский государственный университет, Томск
Сведения о численности
земноводных и пресмыкающихся крайне скудны. Обычно этих животных учитывают на
ограниченных площадях, поэтому представления о распределении в целом по
регионам нет. Западная Сибирь в этом отношении изучена лучше других территорий
России (Куранова, настоящий сборник), по крайней мере,
по земноводным, а Томская и Новосибирская области – и по пресмыкающимся. Это
позволило охарактеризовать и классифицировать население указанных классов
животных, оценить силу связи их сообществ с неоднородностью среды рассматриваемых
областей.
Материалы и методы
Материалы, послужившие
основой для данного сообщения, собраны в 1975-80, 82,85,91-93, 1999-2003 гг.
Учётами охвачены все преобладающие по площади местообитания обеих областей.
Земноводных учитывали с помощью пятидесятиметровых канавок (или заборчиков из
полиэтиленовой плёнки) с пятью цилиндрами, залитыми, как правило, на четверть
четырёхпроцентным раствором формальдегида. Небольшая часть материалов по этой
группе животных собрана при проведении учётов на площадках и маршрутах.
Пресмыкающихся учитывали в основном на маршрутах и реже на площадках. Все результаты
учёта пересчитаны на 1 км2 исходя из ширины
и длины трансекта или площади площадок. Относительные
данные уловов в цилиндры сначала пересчитаны на 100 цилиндро/суток,
а затем на 1 км2 по соотношению
относительных и абсолютных показателей. Поскольку результаты маршрутных учётов
земноводных при высокой активности в 231 раз выше уловов в канавки в пересчёте
на 100 ц/с, все относительные данные по земноводным пропорционально
увеличены. Пресмыкающихся учитывали, как правило, в начале лета (май-июнь), а
земноводных – обычно во второй половине лета (16 июля – 30 августа). При расчёте
среднелетних показателей значения по пресмыкающимся увеличены
в 1,5 раза, поскольку во второй половине лета их обилие возрастается
за счёт размножения примерно вдвое (Приклонский,
Самарина, 1985; Борисович и др., 2002). За показатели обилия земноводных в
первой половине лета приняты значения без учёта сеголеток, а во второй –
результаты реальных отловов в это время, вместе с сеголетками. Для расчёта среднелетнего обилия и те, и другие данные усреднены с
равным весом. После этого все показатели еще раз усреднены по выделам фрагмента
карты «Растительность Западно-Сибирской равнины» (1976, М 1:1500000), охватывающего
территорию Томской и Новосибирской областей. В том случае, если информация по численности
одной из групп животных отсутствовала, использовались данные по близким типам растительных
формаций или они объединялись в один выдел.
Всего нами проанализирована
информация о 377 вариантах населения земноводных и 213 вариантах сообществ пресмыкающихся. В результате усреднения по выделам карты
и объединения части их для дальнейшего анализа получено 55 средних с
показателями обилия всех видов обоих классов позвоночных. При расчёте средних использованы
данные, собранные не только авторами сообщения, но и другими вкладчиками банка
данных лаборатории зоологического мониторинга ИСиЭЖ
СО РАН. В связи с этим, авторы искренне признательны Л.Г.Вартапетову, О.В.Перковской,
Б.Н.Фомину, О.А.Кречетовой, А.А.Соктину, С.В.Патракову, Е.Л.Шору,
Т.И.Казанцевой, С.А.Вицман, О.Ю.Тютенькову,
И.В.Лалетину, О.А. Полушиной, О.Н. Чернышовой,
А.М.Панфилову, И.В.Лукьяновой, И.Г.Тарасову, М.В.Путько,
О.В.Бахтиновой,
И.Е.Евдокимовой, А.В. Андреевских и А.Д. Дубовику за предоставленное право
использовать их материалы и помощь в проведении учётов. Кроме того,
использованы литературные сведения (Григорьев, 1972,1981; Попоудина,
1976).
По рассчитанным,
как описано выше, средним вычислены коэффициенты сходства Жаккара
(Jaccard, 1902) для количественных признаков (Наумов, 1964). По матрице
этих коэффициентов одним из методов кластерного анализа проведена классификация
населения (Трофимов, Равкин, 1980). По итогам анализа
и интерпретации полученного разбиения составлена обобщённая концепция
неоднородности населения земноводных и пресмыкающихся указанных областей, т.е.
формализованное разделение идеализировано (Равкин и
др., 2001). Затем все имеющиеся данные в виде средних
по выделам карты разнесены по таксонам классификации и усреднены. По этим
средним значениям для всех видов и составлены характеристики типов населения. Описание
распределения земноводных и пресмыкающихся проведено по обилию, рассчитанному по
выделам карты растительности. Классификация видов по сходству распределения
выполнена тем же методом и по тем же показателям, но после их перевода в
десятичные логарифмы. Этот перевод выравнивает оценки распределения, что
способствует классифицированию по сходству в размещении по территории. Методы и
принципы классификации здесь не приводятся, поскольку описаны ранее (Равкин и др., 1994а).
Из-за значительной
идеализации представлений, сформированных по результатам формализованной
классификации и усреднения проб, нередко непохожих друг на друга, но
относящихся к выделенным типам согласно интерпретации разбиения, построенный
структурный граф имел явно неудовлетворительное объяснение. Поэтому граф,
приведённый в статье, построен по коэффициентам сходства, рассчитанным по
средним для типов показателям обилия, а не средним коэффициентам сходства между
вариантами, отнесёнными к тем или иным типам, как это делается обычно. Следует
отметить, что земноводных, за исключением озёрной лягушки, подсчитывали только
на суше, т.е. пребывание в водоёмах и реках других видов лягушек, обыкновенного
тритона и углозуба во время икрометания и личиночного
развития не учитывалось.
Выполненные классификации
и граф сходства использованы при составлении карты населения земноводных и пресмыкающихся
и уточнения списка основных факторов среды, определяющих неоднородность их сообществ
в указанных областях. Для оценки силы и общности неоднородности населения и
среды применена линейная качественная аппроксимация – качественный
аналог регрессионной модели (Равкин и др., 1978).
Повидовой обзор
Земноводные
Сибирский углозуб[1] (Salamandrella keyserlingii)
В средней тайге
сибирского углозуба больше всего в
темнохвойно-мелколиственных лесах (1,5), вдвое меньше на низкорослых рямах верховых болот, в сосняках, посёлках и совсем мало
встреч приходится на мелколиственные леса (0,05). В южной тайге максимальные
значения свойственны темнохвойной тайге (1,1), почти вдвое меньше углозуба на внепойменных облесённых низинных болотах (0,7) и примерно в 5 раз – на
гарях и вырубках, шелкопрядниках, в елово-кедровой
тайге и пойменных ивняках в сочетании с лугами. В 10 раз меньшие показатели
характерны для открытых внепойменных низинных болот и
совсем мало ловили его на грядово-мочажинно-озёрных
верховых болотах и в посёлках (0,05 и 0,01).
В подтаёжных лесах углозуб встречен на займищах
(0,3), в берёзово-осиновых лесах и сосняках (по 0,1). В лесостепи отмечен в полях-перелесках (0,2), берёзово-осиновых лесах
(0,05) и посёлках (0,01). В степной зоне углозуб не обнаружен.
На Салаире его ловили только в лесах –
берёзово-сосновых и берёзово-осиновых (по 0,05).
Итак, в лесной зоне в
пределах Томской и Новосибирской областей сибирский углозуб предпочитает темнохвойные и
темнохвойно-мелколиственные леса и лишь при их отсутствии – леса с иным
составом лесообразующих пород или евтрофные болота. В
лесостепи в большем количестве он встречается в колках среди полей, где нередки
мелкие водоёмы, в которых углозубы размножаются.
Верховые и открытые низинные болота, а также поймы крупных рек и черневую низкогорную
тайгу углозуб, как правило, избегает.
Обыкновенный тритон (Triturus vulgaris)
Встречен в подтаёжных
берёзово-осиновых и сосновых лесах, в том числе на останцах в пойме Оби (по
0,2), а также в лесостепи во внепойменных лугах и
мелколиственных лесах (0,1 и 0,08).
Серая жаба (Bufo bufo)
В средней тайге больше
всего серой жабы отмечено в темнохвойно-мелколиственных лесах (8) и в 1,5 раза
меньше в низкорослых рямах верховых болот, в сосняках
и посёлках. Минимальные значения свойственны берёзово-осиновым лесам (2). В
южной тайге, так же как в средней, максимальное обилие свойственно
темнохвойно-мелколиственным лесам (3,5), смешанным полузаболоченным
лесам, низкорослым рямам (по 3) и темнохвойной тайге
(2,7). В 2-3 раза меньше её на гарях и в мелколиственных лесах по долинам
притоков крупных рек. В 6-7 раз меньшие значения характерны для сосняков и полей-перелесков,
а минимальные значения – для елово-кедровой тайги и посёлков (0,2 и 0,1).
В подтаёжных лесах серой
жабы больше всего в низкорослых рямах и сосняках
(примерно 1750 и 600), но эти показатели явно завышены из-за проведения учётов
в непосредственной близости от водоёмов выплода во
время выхода сеголеток. В берёзово-осиновых лесах её обилие достигает 300 тыс.
особей/км2, а в полях-перелесках, на
низинных пойменных болотах и в парках оно значительно меньше (86; 69 и 46). В
северной лесостепи больше всего серой жабы отмечено в берёзово-осиновых лесах и
полях-перелесках (6 и 4). Минимальные значения свойственны соснякам и посёлкам
(0,6 и 0,2). В южной лесостепи и степной зоне в исследованных областях она нами
не встречена.
В салаирской
черневой тайге и на гарях в среднем на 1 км2
держится около 700 особей, и существенно меньше серой жабы в берёзово-сосновых
и берёзово-осиновых лесах (примерно 460 и 150 особей).
Таким образом, в
рассматриваемых областях серая жаба в таёжных подзонах
и на Салаире предпочитает темнохвойно-мелколиственные
леса, а в подтаёжных лесах, где этих местообитаний почти нет, – рямы и сосняки. В северной лесостепи зона предпочтения из
имеющегося набора местообитаний из-за общей сухости смещается в мелколиственные
леса и колки среди полей.
Зелёная жаба (Bufo viridis)
Нередка под
Новосибирском, куда завезена с сельскохозяйственной продукцией из Средней Азии
(Золотаренко, 1985).
Остромордая лягушка (Rana arvalis)
В средней тайге больше
всего этой лягушки в пойменных лугах (20), примерно вдвое меньше на низкорослых
рямах верховых болот (12), втрое – в посёлках,
вчетверо – в мелколиственных лесах и в 6-7 раз – в темнохвойно-мелколиственных
и сосновых лесах. По южной тайге максимальные значения свойственны пойменным
лесам (536) и в 6 раз меньшие – пойменным болотам и сорам.
На низкорослых рямах верховых болот и внепойменным низинных болотах остромордой лягушки меньше в
16-17 раз. Для пойменных лугов и мелколиственных внепойменных
лесов значения уменьшаются в 32 и 35 раз, а для вырубок, гарей и шелкопрядников в 131
раз. В темнохвойной тайге, сосняках, полях-перелесках и посёлках показатели
обилия этой лягушки меньше в 160-170 раз. Совсем немного её в елово-кедровой
тайге и смешанных лесах долин притоков крупных рек (1,6–1,9) и меньше всего на грядово-мочажинно-озёрных верховых болотах (0,9).
В подтаёжных лесах больше
всего остромордой лягушки на внепойменных низинных
болотах (139) и меньше на таких же пойменных болотах (в четыре раза) и в
пойменных лугах, ивняках (в пять раз). В 11 раз меньше этой лягушки в парках
городов. В 24 раза меньшие значения характерны для займищ и мелколиственных
лесов и в 54 раза – для посёлков. Обилие остромордой лягушки близко к минимуму
в сосняках (1,1), на низкорослых рямах верховых болот
и в полях, чередующихся с перелесками (0,8 и 0,7).
В северной лесостепи остромордая
лягушка в наибольшем количестве держится на внепойменных
облесённых низинных и переходных болотах (50) и в пойменных
ивняках (30). Примерно в 3,5 раза меньше её на низинных пойменных болотах и в
12 раз – в полях-перелесках. Ещё меньше этой лягушки в сосняках и
осиново-берёзовых лесах (1,9 и 1,7), а особенно в посёлках и городах диффузного
типа (0,5 и 0,1). В южной лесостепи максимальное обилие свойственно лугам (67)
и на порядок меньше остромордой лягушки в мелколиственных лесах и на займищах.
Немного её в остепнённых лугах и посёлках (5 и 1,9),
а также в полях (1,4). В степной зоне в пределах рассматриваемых областей
пойменные ландшафты не представлены, поэтому наибольшее обилие остромордой
лягушки отмечено на займищах и лугах (14 и 8). Минимальные значения свойственны
полям и посёлкам (7 и 4).
В предгорно-низкогорных салаирских ландшафтах наибольшее обилие остромордой лягушки
отмечено в берёзово-сосновых лесах (14). Меньше её берёзово-осиновых,
на вырубках и в черневой тайге (5, 3 и 0,5).
Таким образом,
остромордая лягушка предпочитает внепойменные открытые
низинные болота и поймы крупных рек, а особенно – зарастающие пойменные соры в южной тайге. От подтаёжных лесов до южной лесостепи
в число предпочитаемых местообитаний входят и облесённые
низинные болота. В лесостепи и степи это, кроме того, осоково-тростниковые
болота и сплавины пресных озёр. В незначительном количестве встречается остромордая
лягушка в сухих сосняках и на участках под пологом темнохвойных пород, если
рядом или внутри нет низинных болот. То же наблюдается и на обширных южнотаёжных грядово-мочажинно-озёрных
болотах с минимальной трофностью и высокой
кислотностью водоёмов, а также на сплавинах и островах среди солёных озёр. В
средней и южной тайге она предпочитает пойменные местообитания, а южнее –
болота.
Сибирская лягушка (Rana amurensis)
В средней тайге отмечена на притеррасном пойменном низинном болоте и в
расположенном рядом посёлке (4 и 0,06). В южной тайге встречалась только в поймах:
в наибольшем количестве в лесах, меньше – в лугах и на сорах
(280; 10 и 5) и в рядом расположенных посёлках (0,5). В подтаёжных лесах её
больше всего ловили на пойменных низинных болотах (4) и существенно меньше в
таких же болотах на террасах, а также в приречных берёзово-осиновых лесах (0,2
и 0,6). В лесостепи сибирскую лягушку встречали на займищах и в прилежащих к
ним лугах (0,5 и 0,1).
Таким образом, сибирская
лягушка в средней и южной тайге предпочитает пойменные низинные болота, меньше
– луга. За пределы пойм почти не выходит. В подтаёжных лесах вне пойм
встречается чаще, но тоже предпочитает низинные болота и луга. В лесостепи
больше всего её на сплавинах пресных озёр, меньше – на осоково-тростниковых
болотах.
Озёрная лягушка (Rana ridibunda)
Завезена при рыборазведении и случайных
выпусках после лабораторных занятий (Малков, Яковлев, 1985; Куранова,
настоящий сборник). Встречается на мелководьях и берегах Обского водохранилища,
прилежащих водоёмах (60) и в озёрах в центре Томска (примерно 2).
Пресмыкающиеся
Прыткая ящерица (Lacerta agilis)
Среднюю тайгу обычно
включают в ареал прыткой ящерицы (Банников и др., 1977). Однако нами в этой подзоне она не встречена, хотя эту ящерицу ловили под
Ханты-Мансийском и вдоль железной дороги до Ноябрьска
(Гашев, Лаврентьев, 2003). В южной тайге её отмечали
только в пойменных лесах (1,2). В подтаёжных лесах больше всего этой ящерицы в
городах диффузного типа и парках на месте сосновых и берёзово-сосновых лесов (3,5
и 2,1), а также в этих исходных насаждениях (1,6). Существенно меньше её на низкорослых
рямах верховых болот (0,9), в берёзово-осиновых лесах
(0,3), а также в полях-перелесках и пойменных лугах, чередующихся с болотами
(по 0,2). В северной лесостепи больше всего этой ящерицы во внепойменных
берёзово-осиновых лесах и на облесённых низинных
болотах (примерно по 0,5). Вдвое меньше её в сосновых и берёзово-сосновых лесах
и втрое – в посёлках. Минимальное обилие свойственно полям-перелескам и пойменным
ивнякам (по 0,1 и 0,02). В южной лесостепи её видели только в посёлках (0,01),
а в колочной степи – в перелесках среди лугов и полей
(Попоудина, 1976).
Итак, прыткая ящерица
встречается в основном в подтаёжных лесах и в значительно меньшем количестве – в
северной лесостепи, а особенно в таёжных подзонах и колочной степи. В подтаёжных лесах предпочитает сосновые и
берёзово-сосновые леса, а в северной лесостепи – берёзово-осиновые, но особенно
широкие просеки, склоны каналов и мелиоративных валов, посадки, обочины дорог и
огороды.
Живородящая ящерица (Zootoca vivipara)
В средней тайге больше
всего этой ящерицы держится на низкорослых рямах
верховых болот (0,5), вдвое меньше – в берёзово-осиновых лесах, а минимальные
показатели характерны темнохвойно-мелколиственным (0,08), сосновым и
берёзово-сосновым лесам (0,04). В южной тайге её больше всего в посёлках и в среднем
в полях-перелесках (1,2 и 2,2). В полузаболоченных
смешанных лесах и пойменных ивняках, чередующихся с лугами, её обилие
колеблется от 0,2 до 0,7 тысяч особей/км2.
На гарях и вырубках, в сосновых лесах и смешанных насаждениях в долинах притоков
крупных рек на ту же площадь приходится 0,4 и по 0,3 тысячи особей, так же как
в елово-кедровой тайге и на низинных болотах (0,3 и 0,2). Минимальные значения
свойственны грядово-мочажинно-озёрным верховым болотам
(0,02).
В подтаёжных лесах больше
всего живородящей ящерицы держится в сосняках и берёзово-сосновых лесах (3,3). Вдвое
меньше её на низкорослых рямах верховых болот (1,4) и
примерно вчетверо – в берёзово-осиновых лесах, полях-перелесках и на внепойменных низинных болотах (0,3–0,4). Существенное снижение
обилия этой ящерицы отмечено в посёлках, на займищах (0,1 и 0,07) и особенно на
низинных пойменных болотах (0,02). В северной лесостепи больше всего её
насчитывали в посёлках (4,6) и полях-перелесках (2) и втрое меньше – в
берёзово-осиновых лесах (1,3) и почти в 10 раз – на облесённых
низинных и переходных болотах (0,6). Минимальные значения свойственны соснякам
и берёзово-сосновым лесам (0,2). В южной лесостепи в целом её обилие ещё ниже
(в берёзово-осиновых лесах и на лугах по 0,2; на займищах и в полях-перелесках по
0,06). В степной зоне эта ящерица не отмечена.
В предгорно-низкогорных
ландшафтах Салаира больше всего живородящей ящерицы
на вырубках (0,6), меньше в берёзово-сосновых лесах (0,4) и вчетверо меньше в
берёзово-осиновых. Минимальная встречаемость отмечена в черневой
осиново-пихтовой тайге (примерно 0,07).
Итак, в
Томской и Новосибирской областях живородящая ящерица предпочитает: в средней
тайге низкорослые рямы верховых болот, в южной тайге
– перелески среди полей, а среди подтаёжных лесов – сосновые и берёзово-сосновые
и в меньшей степени низкорослые рямы. В северной лесостепи, если не
считать посёлков, её больше всего в полях-перелесках и мелколиственных лесах, а
в южной лесостепи – в берёзово-сосновых лесах. Таким образом, у живородящей
ящерицы явно прослеживается смена стаций. В средней тайге она занимает наиболее
прогреваемые, не затапливаемые в половодья местообитания, в южной тайге – колки
и перелески, т.е. уже облесённые, но хорошо прогреваемые
на опушках, местообитания. В подтаёжных лесах это сосновые и берёзово-сосновые
хорошо прогреваемые леса. В северной лесостепи это вновь поля-перелески, а в
южной – мелколиственные леса. То есть в северной части
Томской области она выходит из под полога леса, а в южной – предпочитает опушки.
В подтаёжных лесах с увеличением общей теплообеспеченности зона предпочтения
этой ящерицы смещается в леса, но самые тёплые. В северной лесостепи снова
предпочитаются колки с опушками, видимо, за счёт возможности терморегуляции при
перемещении на опушки и ухода из них. В южной лесостепи оптимальными для неё
становятся мелколиственные леса.
Обыкновенный уж (Natrix natrix)
В средней тайге не
встречается, в южной – больше
всего ужа насчитывали в посёлках (1,2) и изредка видели его в пойменных лугах и
низинных болотах (по 0,002). В подтаёжных лесах, наоборот, больше всего ужа на
пойменных лугах (1), мало в берёзово-осиновых лесах и на пойменных болотах (0,09
и 0,03). В северной лесостепи максимальное обилие ужа отмечено на облесённых низинных болотах (0,3), значительно меньшее – в
берёзово-осиновых, сосновых и берёзово-сосновых лесах (по 0,5) и минимальное –
в полях-перелесках (0,02). В южной лесостепи и степной зоне нам не встречался.
Обыкновенная гадюка (Vipera berus)
В средней тайге больше
всего гадюки отмечено в посёлках и в берёзово-осиновых лесах (по 0,06) и вдвое
меньше на низкорослых рямах верховых болот. В южной
тайге максимальные значения характерны для полей-перелесков, верховых болот и
смешанных лесов в долинах притоков крупных рек (0,5–0,7). В два раза меньше её
в елово-кедровой тайге (0,3) и значительно меньше в берёзово-осиновых лесах, на
вырубках, гарях и шелкопрядниках (по 1). Совсем
немного этой змеи в сосняках и берёзово-сосновых лесах (0,05–0,06), на низинных
болотах (0,03) и в пойменных лугах и ивняках (по 0,02).
В
подтаёжных лесах, так же как в таёжных подзонах, её больше
всего в низкорослых рямах (0,2). Почти вдвое меньше гадюки в
берёзово-осиновых лесах (0,08) и совсем немного – на низинных пойменных
болотах, в полях-перелесках и сосняках (по 0,02). В северной лесостепи
максимальное обилие характерно для облесённых
низинных и переходных болот (0,2) и вдвое меньшее – для мелколиственных лесов
(0,1). Очень мало её в полях-перелесках (0,003). В южной лесостепи гадюки
совсем немного (в лугах – 0,02, в полях – 0,004). В степной зоне нами не отмечена.
Обыкновенный щитомордник (Gloydius halys)
Встречен М.В.Пестовым
(настоящий сборник) на скалах по крутым берегам среднего течения реки Бердь, в пределах лесостепной зоны.
Классификация земноводных и
пресмыкающихся
по сходству распределения
При классификации видов
по сходству распределения выделено 6 типов преференции. Здесь следует
напомнить, что употребляемый нами в классификации термин «предпочитающие» не
совсем точен. Принцип объединения на самом деле соответствует понятию «имеющие
максимальное перекрытие в зонах предпочтения». Иными словами, если в таксоне
классификации больше одного вида, то перечень местообитаний соответствует
совместно предпочитаемым биотопам, т.е. перечисленные местообитания не обязательно
самые излюбленные. Это лишь те биотопы, которые входят в список местообитаний,
предпочитаемых всеми и в среднем видами
(зона совместной преференции). Если упомянут лишь один вид, то в списки таких биотопов,
согласно иерархии таксонов, входят все перечисленные местообитания. При этом
следует учитывать, что, как при всякой классификации, мы в данном случае
проигрываем за счёт огрубления в описании уникальности распределения видов, но
выигрываем, выявляя его наиболее общие черты, получая представления о факторах
среды, в наибольшей степени определяющих распределение животных на конкретной
территории в описываемое время.
В результате составлена
следующая иерархическая классификация.
Виды, предпочитающие:
1 – подтаёжные леса, рямы и, кроме того, –
1.1 – смешанные полузаболоченные южнотаёжные леса,
а также
1.1.1
– леса и низкорослые рямы верховых болот средней
тайги (углозуб),
1.1.2
– перелески среди полей и верховые болота южной тайги (гадюка),
1.1.3
– перелески среди полей в лесостепи (серая жаба, живородящая ящерица),
1.1.4
– низинные болота и поймы крупных рек средней и южной тайги (остромордая
лягушка);
1.2 –
подтаёжные и лесостепные посёлки, города диффузного типа и парки (прыткая
ящерица);
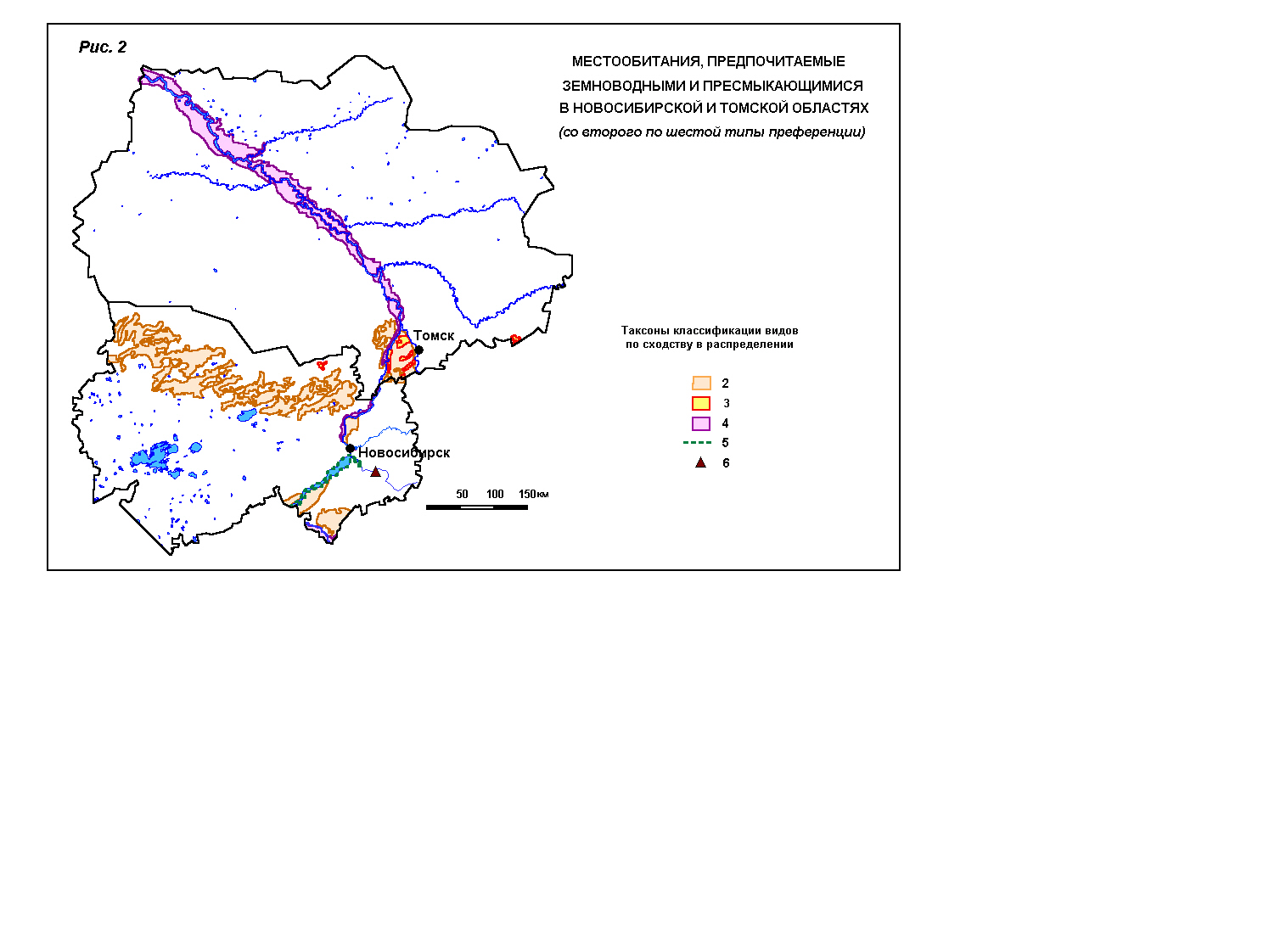
2 –
подтаёжные и лесостепные леса (тритон);
3 –
лесостепные облесённые низинные болота (зелёная жаба)
и, кроме того, сосновые подтаёжные леса (уж);
4 –
поймы крупных рек (сибирская лягушка);
5 –
мелководья и берега Новосибирского водохранилища и прилежащих водоёмов (озёрная
лягушка);
6 – скалы по крутым берегам в среднем
течении р.Бердь
(щитомордник).
Таким образом, лишь два
типа предпочтения имеют иерархическое деление, а остальные представлены отдельными
видами с уникальным распределением. На картосхемах 1 и 2 цветовыми линиями и
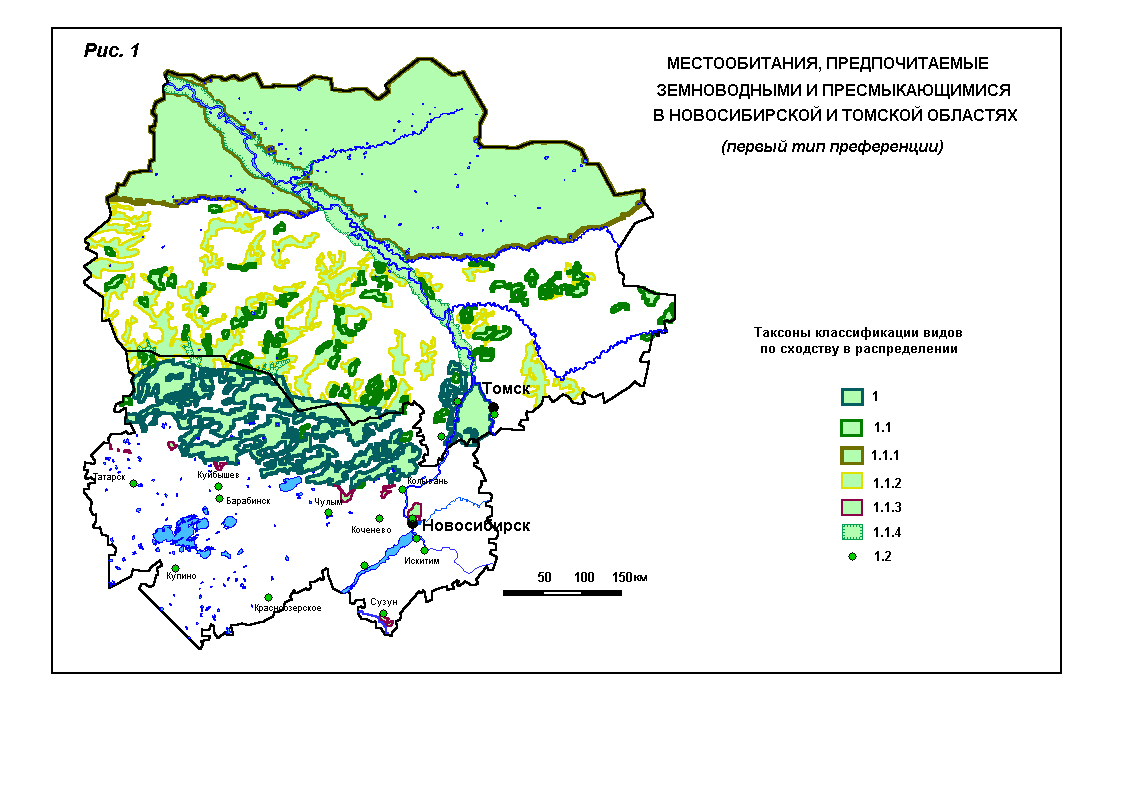
фоном отображены предпочитаемые местообитания[2].
Так, на рис. 1 зелёным цветом обозначены все местобитания, предпочитаемые видами, которые отнесены к
первому типу предпочтения. Синим цветом обведены те
местообитания, которые предпочитаются в целом видами этого типа. Кроме этих
биотопов разные виды (в одном случае пара видов) предпочитают еще специфичные
для них местообитания. Они обведены на карте линиями разных цветов. Так,
например, углозуб отнесен к
первому типу, первому подтипу (1.1) и первому классу распределения (1.1.1.).
Таким образом, в зону его преференции входят подтаёжные леса и рямы (1 – синяя обводка), смешанные полузаболоченные
южнотаёжные леса (1.1 – зелёная обводка), а также
леса и низкорослые рямы верховых болот средней тайги
(1.1.1 – коричневая обводка). Остромордая лягушка предпочитает те же местообитания,
помеченные индексами 1 и 1.1, но её распределение отличается тем, что в список излюбленных
местообитаний входят не среднетаежные леса и верховые болота, а низинные болота
и поймы крупных рек средней и южной тайги (1.1.4 – обводка тонкими зелеными чертами).
Информативность этих
представлений такова: по исходному среднему обилию классификация учитывает
всего 8% дисперсии матрицы нецентрированных коэффициентов
корреляции распределения видов и 67% после логарифмирования этих показателей
(коэффициенты корреляции 0,28 и 0,82). Население
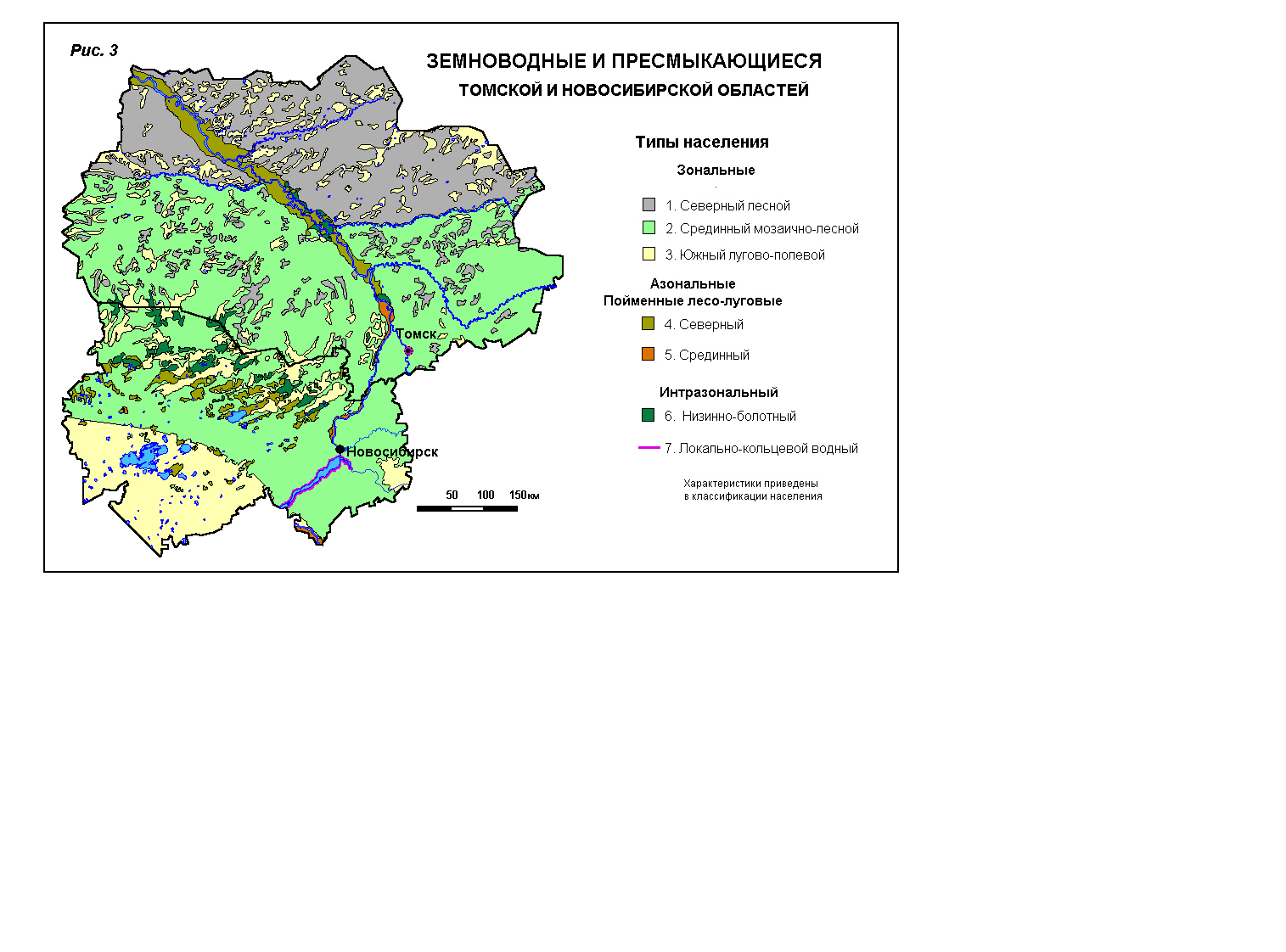
земноводных и пресмыкающихся Для Томской и
Новосибирской областей характерны семь типов сообществ земноводных и
пресмыкающихся. Из них три зональных – северный, срединный,
южный и два азональных (пойменных) – северный и
срединный. Из двух оставшихся типов один интразональный (диффузный), т.е. распространённый
мелкими включениями по территории, занимаемой зональными типами. Последний тип
населения, названный локально-кольцевым, не связан с зональной изменчивостью
климата и охватывает Обское водохранилище по берегам и мелководьям.
Характеристики сообществ приведены ниже. Классификация населения земноводных
и пресмыкающихся Томской и Новосибирской областей Типы населения Зональные 1. Северный лесной (лесов средней тайги с проникновением в южную темнохвойную тайгу). Серая
жаба 3745[3],
остромордая лягушка 3665, сибирский углозуб 737, живородящая ящерица 113, гадюка 65, сибирская
лягушка 10; всего 8289 / 6 видов. 2. Срединный
мозаично-лесной (нетемнохвойных лесов, вырубок, гарей, шелкопрядников,
полей-перелесков южной тайги, лесов – подтаёжных и северной лесостепи с
проникновением на рямы, в том числе среднетаёжные, и нечерневых салаирских лесов). Серая жаба 82237, остромордая лягушка
5467, живородящая и прыткая ящерицы 910 и 388, углозуб
99, гадюка 94, сибирская лягушка 80, озёрная лягушка 58, уж 55, тритон 31;
88961 / 10. 3. Южный лугово-полевой (полей, лугов, степей южной лесостепи и степной зоны с
проникновением на грядово-мочажинно-озёрные таёжные
верховые болота и в салаирскую черневую тайгу).
Остромордая лягушка 10587, серая жаба 86, углозуб 54,
гадюка 58, живородящая и прыткая ящерицы 38 и 19, тритон 13, сибирская лягушка
11; 10803 / 8. Азональные (пойменные лесо-луговые) 4. Северный (в пределах таёжных подзон с проникновением на
займища от подтаёжных лесов до степи). Остромордая и сибирская лягушки 99996 и
49252, прыткая и живородящая ящерицы 216 и 129, углозуб
92, гадюка и уж 8 и 0,3; 149337 / 7. 5. Срединный (подтаёжно-лесостепной с проникновением на лесостепные
осоково-вейниковые внепойменные болота). Остромордая
и сибирская лягушки 31359 и 970, прыткая и живородящая ящерицы 207 и 124,
зелёная жаба 92, уж и гадюка 73 и 59, серая жаба 17; 32446 / 8. 6. Интразональный низинно-болотный (южной тайги и подтаёжных лесов).
Остромордая и сибирская лягушки 75490 и 1398, серая жаба 875, уж 245,
живородящая ящерица 152, углозуб 29, гадюка 8; 78196
/ 7. 7. Локально-кольцевой водный (мелководий и берегов Обского водохранилища с проникновением на прилежащие к нему водоёмы и озера в Томске). Озёрная
лягушка 59832. Первый
из зональных типов (лесной) занимает большую часть средней тайги, второй
(мозаично–лесной) – южной тайги, подтаёжных лесов и северной лесостепи, третий
(лугово–полевой) тип населения – южной лесостепи и
степи. Азональные (лесо–луговые) типы распространены по лугам и лесам пойм
крупных рек, соответственно, северный – в пределах таёжных подзон
(средней и южной), а срединный тип населения – таких же пойменных местообитаний
в пределах подтаёжных лесов и северной лесостепи. Интразональный тип сообществ свойствен низинным и переходным болотам южной тайги и
подтаёжных лесов. Локально-кольцевой, как уже отмечено, связан с лесостепным
Обским водохранилищем (рис. 3).
Северный
зональный тип занимает в основном территорию внепойменных
лесов и вырубок средней тайги, срединный – нетемнохвойные
и нечерневые леса (светлохвойные, мелколиственные и
хвойно-лиственные, кроме осиново-пихтовых), а также вырубки, гари, шелкопрядники, поля-перелески и рямы
от южной тайги до северной лесостепи и на Салаире. Южный зональный тип населения
свойствен, в основном, полям, лугам и луговым степям в пределах южной лесостепи
и степной зоны. Сообщества всех посёлков и городских окраин входят в те типы
населения, на территории которых они расположены. Про приуроченность незональных
типов уже говорилось. Для всех типов населения,
помимо их основного ареала, характерно проникновение на территории, занятые
преимущественно другими типами в виде изолированных анклавов. Так, северный тип
(среднетаёжный лесной) проникает в южную тайгу по участкам темнохвойных лесов.
Срединный мозаично-лесной тип населения свойствен среднетаёжным
рямам верховых болот. Южный зональный тип сообществ
характерен ещё и для грядово-мочажинно-озёрных
комплексов верховых таёжных болот и черневой (осиново-пихтовой) салаирской тайги. Азональный
северный тип сообществ занимает, кроме пойм, ещё и тростниковые займища от
подтаёжных лесов до степи, а срединный пойменный – лесостепные внепойменные осоково-вейниковые болота. Интразональный
диффузный тип населения весь размещён в анклавах, как в поймах, так и вне их
территории. Локально-кольцевой тип проникает на мелкие водоёмы, прилежащие к
Новосибирскому водохранилищу. Судя по плотности
населения, оптимальные и субоптимальные условия
обитания свойственны территории, занимаемой северным азональным
и срединным мозаично-лесным типами (около 149 и 89 тысяч особей/км2). Интразональный и локально–кольцевой типы населения
занимают пространства с субпессимальными условиями
существования (около 78 и 60 тысяч). Пессимальные
условия обитания характерны в целом для населения срединного азонального типа, субэкстремальные
– для южного зонального и экстремальные – для северного зонального (примерно 32,
11 и 8 тысяч особей/км2). Общее число видов
колеблется в большинстве типов от 7 до 10 и лишь в экстремальных условиях отмечено
6 видов, а в водном локально-кольцевом типе учитывалась только озёрная лягушка.
Случайные встречи других земноводных и пресмыкающихся здесь во внимание не принимались. Во всех типах населения
суши в число лидеров, т.е. первых трёх по численности видов, всегда входит
остромордая лягушка. В водном типе она занимала бы второе место по
встречаемости, но её не учитывали, т.к. в воду эти лягушки, кроме периода
икрометания, прыгают с берега только при вспугивании. Остромордая
лягушка лишь в двух зональных типах населения – лесном и мозаично-лесном
уступает первое место по обилию серой жабе, которой принадлежит второе место в
оставшемся (южном) зональном типе населения и третье – на низинных болотах южных
подзон лесной зоны. Сибирская лягушка занимает
второе место на этих болотах и в поймах крупных рек, а углозуб
– в среднетаёжном лесном типе. Прыткая ящерица входит
в число лидеров в среднем в азональных типах, живородящая
– в мозаично-лесном, а гадюка – лугово-полевом. Доля
остромордой лягушки в населении всех типов, кроме водного,
колеблется от 5 до 98%, а остальных лидеров от 0 до: серой жабы 93%, сибирской
лягушки 33%, углозуба 9%, а ящериц – прыткой и живородящей
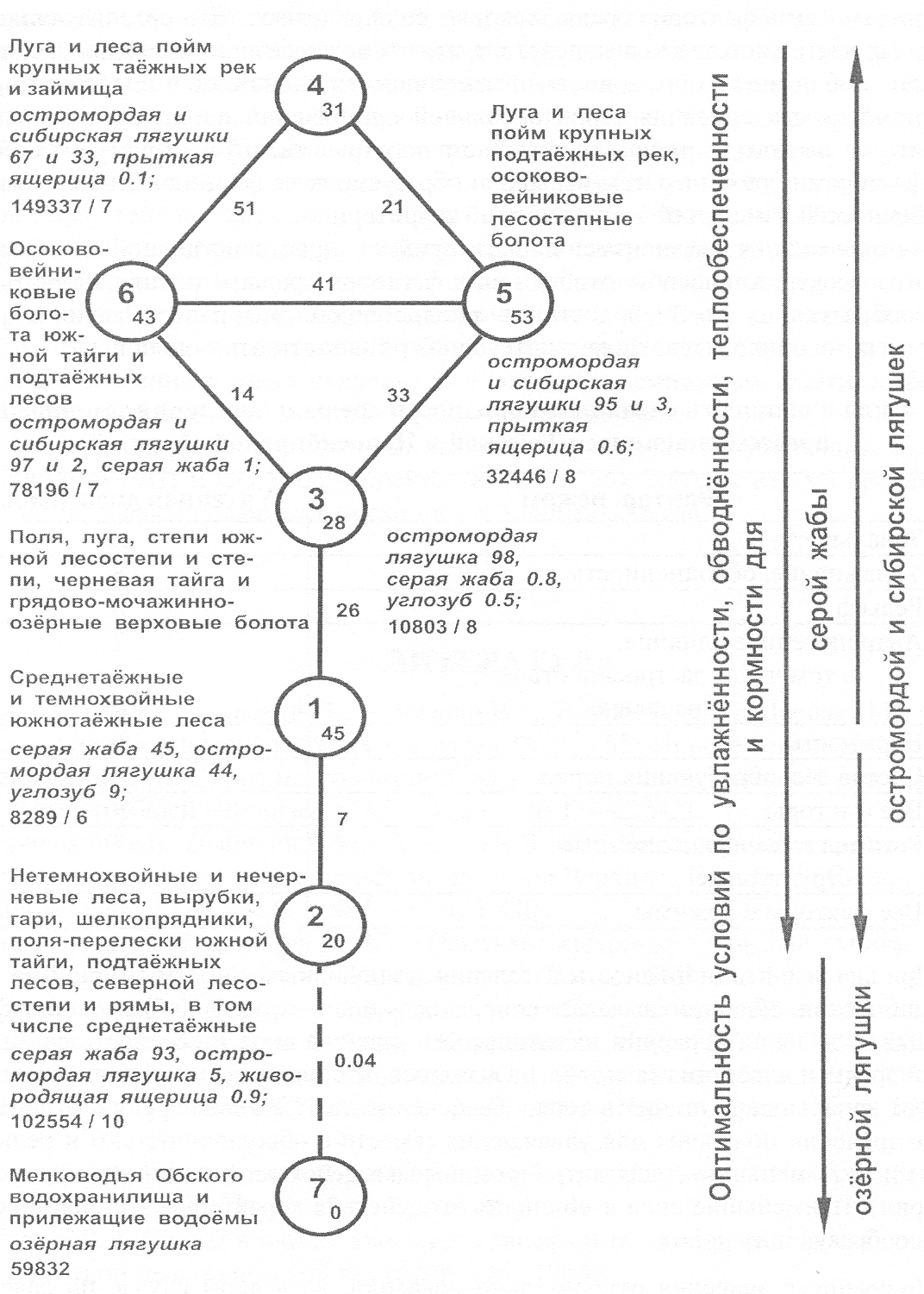
до 1 и 0,5%. Граф, представленный на
рис. 4, иллюстрирует изменения населения в результате интегрального влияния
различий в оптимальности условий обитания по увлажнению, обводнённости,
теплообеспеченности и кормности дифференцированно для
серой жабы; остромордой и сибирской лягушек; озёрной лягушки. При этом направления выявленных изменений на большей части
территории не совпадают у серой жабы и упомянутых видов лягушек, а также у
озёрной и остальных видов лягушек.
Рис. 4.
Пространственно-типологическая структура населения земноводных и пресмыкающихся
Томской и Новосибирской областей. Схема построена по сходству показателей обилия,
усреднённых по типам населения (1-7). Межтиповое
сходство приведено у значимых связей (сплошных линий, прерывистой чертой показана
слабая запороговая связь). Индексом у номера типа
дана внутритиповая общность, рассчитанная по средним
коэффициентам сходства вариантов населения, усреднённых по выделам карты растительности.
У значка с номером типа, помимо перечня занимаемых местообитаний, приведены три
первые по обилию вида с указанием доли каждого из них в населении (%), а также
плотность населения (особей/км2) и число
встреченных видов. В целом, в соответствии со
степенью оптимальности условий существования, плотность населения убывает от
четвёртого типа к первому, а потом возрастает ко второму. Обилие серой жабы
уменьшается в обратном направлении (от второго типа к четвёртому). В этих двух
последних типах доминирует уже серая жаба, а не остромордая лягушка, хотя
обилие последней, так же как серой жабы, возрастает от первого ко второму типу.
Таким образом, за счёт самых различных неблагоприятных факторов среды, по
закону минимума (Либих, по Пианка, 1981) могут формироваться
сходные комплексы земноводных и пресмыкающихся. Неоднородность их населения
формируется, кроме того, по принципам обеднения–обогащения, независимого замещения
(Равкин и др., 1994 б) и смены стаций (Бей-Биенко, 1966). Пространственная
организация населения Под пространственной
организацией животного населения понимается общий характер его территориальной
неоднородности, т.е. его пространственная структура, а также набор и
взаимосвязь факторов среды, которые её определяют. Это
организующая, упорядочивающая часть системы взаимодействия животного населения
и среды, рассматриваемая вместе с общими закономерностями изменчивости сообществ в пространстве. Сложность, громоздкость описания
территориальной организации населения вынуждает рассматривать её дважды –
сначала в основном пространственную структуру сообществ в связи
с факторами среды, а затем иерархию образующих её факторов с повтором некоторых
особенностей изменений в населении по территории. Методы оценки связи
изменчивости среды и пространственной неоднородности животного населения,
способы отображения факторов среды и оценки полноты набора структурообразующих
факторов достаточно подробно описаны ранее (Равкин и
др., 1978), поэтому здесь не приводятся. Ниже даются лишь результаты этих
оценок. При анализе
неоднородности населения земноводных и пресмыкающихся Томской и Новосибирской
областей выявлен список основных структурообразующих факторов среды. Для
выявления иерархии их значимости оценена сила и
общность связи неоднородности среды и населения (см. табл.). Оказалось, что
наибольшее влияние на население оказывают зональные различия в
теплообеспеченности (12% дисперсии). В полтора раза меньшие значения получены
для увлажнения (вместе с обводнённостью) и рельефа (поемности), и чуть меньшие – для антропогенного воздействия
(в основном застроенности территории). Наименьшие
сила и общность воздействия характерны для кормности
и состава лесообразующих пород. Таблица Сила и общность
связи неоднородности среды и населения земноводных и пресмыкающихся Томской и
Новосибирской областей Фактор, режим Учтённая дисперсия, % Зональность 12 Увлажнение, обводнённость 8 Рельеф 8 Антропогенное влияние, 7 в том числе застроенность 7 распашка 0,5 Кормность 6 Состав лесообразующих пород 6 Все факторы 40 Режимы классификационные 4 структурные 10 Все факторы и режимы 47 Полученные значения
относительно невелики, во всяком случае по сравнению с
оценками по птицам и мелким млекопитающим (Равкин,
2002), но в целом, видимо, из-за меньшей скоррелированности
воздействия с перечисленными факторами, можно связать 40% дисперсии населения
(в виде матрицы коэффициентов сходства). Режимные представления, т.е. влияние
неразделимых сочетаний этих факторов, учитывают по классификации сообществ всего 4%, а структурные отображения – 10% дисперсии. В
целом, влияние перечисленных факторов и режимов можно объяснить 47% дисперсии,
что почти вдвое превышает аналогичные оценки по Западно-Сибирской равнине. Заключение Таким образом, в Новосибирской и Томской областях встречено семь видов
земноводных и 5 видов пресмыкающихся, в том числе два вида хвостатых
земноводных (сибирский углозуб и обыкновенный
тритон), два вида жаб (серая и зелёная) и три вида бурых лягушек (остромордая,
сибирская и озёрная). Из пресмыкающихся отмечено два вида ящериц (прыткая и
живородящая) и три вида змей (уж, гадюка и щитомордник). Эти виды формируют
семь типов населения: три зональных, два азональных,
один интразональный и один локально-кольцевой. При этом границы распространения
типов по территории не совпадают с подзональными,
хотя основным структурообразующим фактором среды является зональная
теплообеспеченность. Воздействие других факторов – облесённости, увлажнения (обводнённости),
рельефа (пойменности) и застроенности
примерно в 1,5 раза меньше. В целом, факторами среды удаётся объяснить около
половины дисперсии населения (47%, множественный коэффициент корреляции – 0,63).
В общем невысокая информативность полученных представлений
обусловлена значительным влиянием локальных концентраций особенно у пресмыкающихся
и сеголеток земноводных. Эти особенности не учитываются в выбранном масштабе и
ранге рассмотрения. Исследования, результаты
которых изложены в настоящей публикации, поддержаны Президиумом РАН и СО РАН (проекты
№ 12.2 и 56). Авторы весьма признательны Т.В.Котовой за редактирование карто-схем и ряд ценных советов. Литература Банников А.Г., Даревский И.С., Ищенко В.Г., Рустамов А.К., Щербак Н.Н.
Определитель земноводных и пресмыкающихся фауны СССР. – М.: Просвещение, 1977. Бей-Биенко Г.Я. Смена местообитаний наземными
организмами как биологический принцип // Журн. общей биологии, 1966. – Т. 27. –
№ 1. – С. 5-21. Борисович О.Б., Цыбулин С.М., Торопов К.В. и др. Земноводные и пресмыкающиеся
равнинных и предгорно-низкогорных ландшафтов Верхнего Приобья // Сибирский экологический журнал, 2002. – Т. 9. –
№ 4. – С. 425-440. Гашев С.Н., Лаврентьев С.Ю. Прыткая
ящерица// Красная книга Ханты-Мансийского автономного округа. – Екатеринбург: «Пакрус», 2003. – С. 99. Григорьев О.В. К биологии
сибирского углозуба в
лесостепной зоне Западной Сибири // Зоологические проблемы Сибири. – Новосибирск:
Наука, 1972. – С. 300-301. Григорьев О.В. К экологии
земноводных и пресмыкающихся в пойме Оби // Сукцессии животного населения в
биоценозах поймы реки Оби. – Новосибирск: Наука, 1981. – С. 154-160. Золотаренко Г.С. О
нахождении зелёной жабы в окрестностях Новосибирска // Вопросы герпетологии. –
Л.: Наука, 1985. – С. 80-81. Наумов Р.Л. Птицы в
очагах клещевого энцефалита. Автореф. … канд.дисс. Московский областной
педагогический институт. – М., 1964. Пианка Э. Эволюционная экология. – М.: Мир,
1981. Попоудина А.Д. О питании и размножении ящериц
в южной части лесного Приобья
// Фауна и экология животных Приобья. – Новосибирск:
Наука, 1976. – С. 36-42. Приклонский С.Г., Самарина Б.Ф. Некоторые
параметры популяции обыкновенного ужа в средней полосе // Вопросы герпетологии.
– Л.: Наука, Лениградское отд-ние,
1985. – С. 171. Равкин Е.С., Равкин
Ю.С., Вартапетов Л.Г. и др. Классификация летнего
населения птиц равнин Северной Евразии // Сибирский экологический журнал, 2001.
– Т. 8. – № 6. – С. 741-766. Равкин Ю.С. Пространственно-типологическая
организация животного населения Западно-Сибирской равнины (на примере птиц,
мелких млекопитающих и земноводных) // Зоологический журнал, 2002. – Т. 81. – Вып. 9. – С. 1166-1184. Равкин Ю.С., Вартапетов
Л.Г., Колосова Е.Н. и др. Видовое разнообразие птиц Западно-Сибирской равнины и
общие особенности их летнего распределения // Сибирской экологический журнал,
1994а. – Т. 1. – № 6. – С. 321-335 Равкин Ю.С., Вартапетов
Л.Г., Юдкин В.А. и др. Пространственно-типологическая
структура и организация летнего населения птиц Западно-Сибирской равнины //
Сибирский экологический журнал, 1994 б. – Т. 1. – № 4. – С. 303-320. Равкин Ю.С., Куперштох
В.Л., Трофимов В.А. Пространственная организация населения птиц // В кн.: Ю.С.Равкин . Птицы лесной зоны Приобья. – Новосибирск: Наука, 1978. – С 253-269. Трофимов В.А., Равкин Ю.С. Экспресс-метод оценки связи пространственной
неоднородности животного населения и факторов среды // Количественные методы в
экологии животных. – Л., 1980. – С. 113-115. Яковлев В.А., Малков Н.П.
Новый элемент в фауне амфибий Алтая // Вопросы герпетологии. – Киев: Наукова думка, 1985. – С. 244-245. Jaccard P. Lois de distribution florale
dans la zone alpine. – Bull. Soc.
Vaund. Sci. Nat., 1902. – Vol. 38. – P. 69-130.