УДК 591.9(5-013)+591.526+598.2/9
CЕЗОННЫЕ ОСОБЕННОСТИ
ЗОНАЛЬНЫХ ИЗМЕНЕНИЙ
НАСЕЛЕНИЯ ПТИЦ зАПАДНО-сИБИРСКОЙ РАВНИНЫ
Л.Г. Вартапетов, С.М. Цыбулин, С.П. Миловидов
Институт систематики и экологии животных СО РАН, Новосибирск 630091
В результате многолетнего изучения населения птиц во всех
природных зонах Западно-Сибирской равнины выявлены: преобладающие виды,
суммарное обилие, видовое богатство и биомасса птиц, а также их фаунистический
состав, ярусное распределение и соотношение основных групп потребляемых кормов.
Зональные изменения летнего населения птиц от степной до тундровой зоны
проявляются преимущественно в резкой смене бореальных орнитокомплексов
гипоарктическими при переходе от средней к северной тайге. Обеднение населения,
а также постепенное и последовательное замещение одних видов другими выражены в
меньшей степени. Северотаежные, лесотундровые и даже тундровые летние
орнитокомплексы отличаются от расположенных южнее более высокими показателями
плотности населения и биомассы; большей долей птиц, собирающих корм на земле и
воде; возрастанием доли вегетативных частей растений и позвоночных животных в
рационе птиц; увеличением роли в сообществах транспалеарктов и арктических
видов. Зимой от подтаежных лесов до лесотундры резко снижается суммарное обилие
птиц и количество бореальных видов. На юге Западной Сибири (от южной тайги до
степи) зональные изменения населения птиц менее выражены и более постепенны в
связи с большей давностью и интенсивностью антропогенной трансформации
местообитаний. Они сводятся в основном к смене лесных видов летом птицами
агроценозов и зимой синантропами. При переходе от летнего к зимнему сезону для
западносибирских орнитокомплексов в целом возрастает доля птиц, кормящихся в
кронах и на стволах деревьев, и участие представителей сибирского и
арктического типов фауны, а также становится характерным преобладание семян,
ягод и кормов антропогенного происхождения.
С
середины XIX в.
отмечалась зависимость богатства фауны от теплообеспеченности территории
(Schmarda, 1853) и убывание числа видов животных и растений от экватора к
полюсам (Jakobi, 1919; Pianka, 1966; Kerr, Packer, 1997 и др.). При более
детальном рассмотрении выяснилось, что плотность и видовое богатство населения
животных, в том числе птиц, уменьшается в Европе к северу и югу от
широколиственных лесов или лесостепных дубрав в связи с нарастанием дефицита
тепло- или влагообеспеченности и уменьшением продуктивности экосистем (Гладков,
1958; Гиляров, Чернов, 1974; Чернов, 1975; Базилевич, 1993 и др.).
Плотность
населения птиц лесной зоны Евразии коррелирует не столько с продуктивностью
биоценозов, сколько с продолжительностью безморозного периода (Пузаченко,
1967). Кроме того, для Северной Америки показано уменьшение видового богатства,
разнообразия и выравненности сообществ птиц при изменениях от мягкого, влажного
и стабильного климата к суровому, сухому и нестабильному (Rotenberry, 1978). В
среднесибирском и отчасти западносибирском секторах Евразии прослеживается
значимая корреляция между видовым богатством конкретных фаун и среднеиюльской
температурой воздуха, особенно значимая в тундровой зоне по сравнению с
расположенными южнее (Чернов, 1999). Это хорошо согласуется с результатами
наших детальных исследований зональных изменений летнего населения птиц
Западной Сибири, которые показали преобладающее воздействие широтно-зональных,
в том числе климатических природных факторов по сравнению с
ландшафтно-биотопическими факторами, которые доминируют южнее, в формировании
северотаежных, лесотундровых и тундровых орнитокомплексов (Вартапетов, 1998).
В
умеренных широтах зимняя смертность птиц является ведущим фактором,
определяющим многолетнюю и сезонную динамику их популяций (Паевский, 1999;
Соколов, 1999), поэтому детальные исследования численности и распределения
зимующих птиц представляются нам особенно необходимыми. Зимнее население птиц
лесной зоны Восточной Европы и Сибири становится беднее к северу и востоку.
Причем на этой огромной территории от подтаежных лесов до северной тайги
формируется единый по составу преобладающих видов зимний орнитокомплекс с
преобладанием представителей сибирского типа фауны (Бутьев, 1970; Цыбулин,
1977; Андреев, 1980). Однако зимнее население птиц Западно-Сибирской равнины до недавнего времени оставалось почти
не изученным, что определило необходимость наших специальных и целенаправленных
его исследований. В результате воздействие широтной зональности на формирование
зимних орнитокомплексов оценено как один их наиболее значимых факторов
(Вартапетов и др., 2001). Население птиц Западной Сибири 5-7 месяцев в году
является типично зимним. Поэтому изучение воздействия широтно-зональных
факторов на формирование зимних орнитокомплексов Западной Сибири составляет
основную задачу нашего исследования. Кроме того, появилась возможность
сопоставления с аналогичными результатами, ранее полученными для летнего
населения (Равкин и др., 1994), с целью выявления воздействия сезонной смены природных
условий на зональные изменения населения птиц.
МАТЕРИАЛ И МЕТОДИКА
Птиц
учитывали на постоянных, но не строго фиксированных маршрутах, без ограничения
дальности обнаружения, по методике, разработанной Равкиным (1967). Суть этого
метода сводится к оценке расстояния от учетчика до каждой встреченной птицы
(или группы птиц) и раздельному интервальному пересчету на площадь по группам
заметности и средним дальностям обнаружения. Обоснование этой методики, техника
проведения учетов и обработка данных в последнее время вновь детально
излагаются в работах Равкина с соавторами (1999), а также Ливанова и Равкина
(2001). Учеты проводились преимущественно в феврале с 1975 по 1997 гг. во всех
зонах Западно-Сибирской равнины, за исключением тундровой зоны. Всего обследовано
412 ландшафтных урочищ, в каждом из которых за сезон с учетом проходили по
10 км, а суммарная протяженность учетных маршрутов составила около 7000 км. Кроме того, использованы аналогичные сведения за
летний период, результаты анализа которых были частично опубликованы ранее
(Равкин и др., 1994).
Для расчета усредненных
обобщающих показателей населения птиц по природным зонам и подзонам
использовали соотношения площадей выделов карты "Растительность Западно-Сибирской
равнины" (1976). Для оценки
биомассы и ярусного распределения птиц, а также состава потребляемых ими
кормов, наряду с собственными материалами, использованы сведения из монографии
«Птицы Советского Союза» (1951-1954). В качестве ярусов рассматривались
основные слои или блоки экосистем (Реймерс, 1990), а не только ярусы
растительности. Ярусное распределение видов птиц оценивали по месту сбора корма
с учетом его сезонных изменений. В случаях, когда вид кормится в нескольких
ярусах, принято, что в каждом из них держатся равные доли особей. Несомненно,
что выполненные расчеты приблизительны, но они позволяют судить о
биоценотической роли птиц в различных ландшафтно-зональных условиях. Отнесение
птиц к различным типам фауны проводили по схеме, предложенной Штегманом (1938).
Представления о гипоарктическом и бореальном биогеографическом поясах, а также
о гипоарктическом, бореально-гипоарктическом и бореальном авифаунистических
комплексах заимствованы у Кищинского (1980, 1988). Таксономические названия
птиц приведены по Степаняну (1990), за исключением обыкновенной и пепельной
чечеток (Acanthis flammea, A. hornemanni), которые вслед за Портенко
(1960) рассматриваются как один вид - чечетка (A. flammea).
РЕЗУЛЬТАТЫ
В
летний период соседние зоны и подзоны характеризуются 1-3 общими преобладающими
видами (табл. 1). При этом зональные и подзональные орнитокомплексы
объединяются в 3 группы с существенно различающимся составом этих видов. Первая
группа – лесотундровая, занимающая тундру, лесотундру и северную тайгу,
характеризуется преобладанием птиц редколесий и открытых ландшафтов: желтой
трясогузки (Motacilla flava), овсянки-крошки (Emberiza pusilla) и
чечетки (Acanthis flammea). В этой группе наиболее обособлены тундровые
орнитокомплексы. Причем только в последних, наряду с указанными тремя видами,
общими для тундры, лесотундры и северной тайги, доминируют краснозобый конек (Anthus
cervinus) и подорожник (Calcarius lapponicus). Вторая группа
орнитокомплексов, обитающих в средней, южной тайге и подтаежных лесах, где
наиболее многочисленны настоящие лесные и опушечные виды: вьюрок (Fringilla
montifringilla), буроголовая гаичка (Parus montanus), лесной конек (Anthus
trivialis), белошапочная овсянка (Emberiza leucocephala). Третья
группа – лесостепная, которая занимает лесостепную и степную зону, определяется
преобладанием птиц агроландшафтов – полевого жаворонка (Alauda arvensis)
и грача (Corvus frugilegus).
В
зимний период также прослеживаются постепенные широтные изменения состава
наиболее многочисленных видов (табл. 2). В соседних зонах и подзонах таких
общих видов от 1 до 4. Если летом состав преобладающих видов наиболее близок на
севере равнины (от тундры до северной тайги), где доминируют гипоаркты, то
зимой его общность больше в срединной и южной частях (от средней тайги до
степи), где преобладают лесные и синантропные птицы. При этом орнитокомплексы
объединяются в 4 группы. Первая из них – лесотундровая, характеризуется
абсолютным преобладанием белой куропатки (Lagopus lagopus), которая в
значительном количестве мигрирует из тундровой зоны (Назаров, 1968;
Сыроечковский, Рогачева, 1968; Пиминов, 1985). Вторая группа – северотаежная,
где доминирует сероголовая гаичка (Parus cinctus), наиболее характерный
зимний обитатель северотаежных лесов и редколесий. Третья группа – лесная
распространена от средней тайги до подтаежных лесов. Здесь преобладают
буроголовая гаичка, обыкновенный поползень (Sitta europaea), чечетка и
обыкновенный снегирь (Pyrrhula pyrrhula). Четвертая, лесостепная группа
включает в себя орнитокомплексы лесостепи и степи и характеризуется наиболее
высокой долей синантропных видов: домового и полевого воробьев (Passer
domesticus, P. montanus) и сороки (Pica pica).
Итак, широтно-зональная дифференциация населения птиц и
природных условий в целом на Западно-Сибирской равнине не совпадают. Так, лесостепные
орнитокомплексы наиболее сходны со степными. Летом в лесостепи и степи
доминируют птицы агроценозов, поскольку последние занимают значительно большую
площадь по сравнению со слабо нарушенными ландшафтами. Зимой в обеих зонах
существенно преобладают по численности синантропные виды. Следует учитывать,
что человек заселил степи и лесостепи Западной Сибири существенно раньше, и
плотность его населения здесь гораздо больше, чем в зонах, расположенных
севернее. Поэтому формирование населения птиц на юге Западной Сибири
определяется антропогенным воздействием. Далее к северу, от подтаежных лесов до
средней тайги, в орнитокомплексах преобладают лесные виды птиц. В северной
тайге они сменяются гипоарктическими, которые связаны с кустарниковой и кустарничковой
растительностью больше, чем с лесной, населяют летом все типы ландшафтов и
доминируют также в лесотундре и южной тундре. В зимний период в северной тайге
преобладают как гипоарктические, так и бореальные виды, поэтому северотаежные
орнитокомплексы выделены в качестве отдельной группы.
Суммарное
обилие птиц летом неуклонно уменьшается от степи и лесостепи до средней тайги,
а затем заметно возрастает в северной тайге и несколько снижается в лесотундре
и тундре (рис. 1). Возрастание плотности населения птиц в северной тайге, по
сравнению со средней, отмечалось и ранее в долине Оби (Равкин, 1978), в долине
Енисея (Анзигитова и др., 1979), на таежных междуречьях Западной Сибири
(Вартапетов, 1984) и в Северной Европе (Jarvinen, Sammalisto, 1976; Jarvinen,
Vaisanen, 1980). Теперь о том же свидетельствуют и объединенные материалы для
всей Западно-Сибирской равнины. Среди причин этого увеличения в Западной Сибири
рассматривались такие, как возрастание интенсивности пред- и послегнездовых
перемещений птиц (хотя увеличение прослеживается и по плотности населения в
гнездовый период); высокая численность птиц, гнездящихся также в лесотундре и
северотаежных редкостойных лесах; большая мозаичность местообитаний, особенно
лесоболотных (Вартапетов, 1980, 1984); гетеротрофность северотаежных болот
(Равкин, 1978). Для Северной Европы увеличение численности птиц на
северотаежных болотах по сравнению с расположенными южнее связывается с
оптимальным режимом увлажнения этих болот для водных беспозвоночных,
способствующим их интенсивному размножению. Здесь более развиты кустарники и
кустарнички, которые облегчают кормодобывание и гнездование птиц; а также
больше незаболоченных местообитаний (редколесий, берегов водоемов), пригодных
для гнездования болотных птиц (Jarvinen, Sammalisto, 1976).
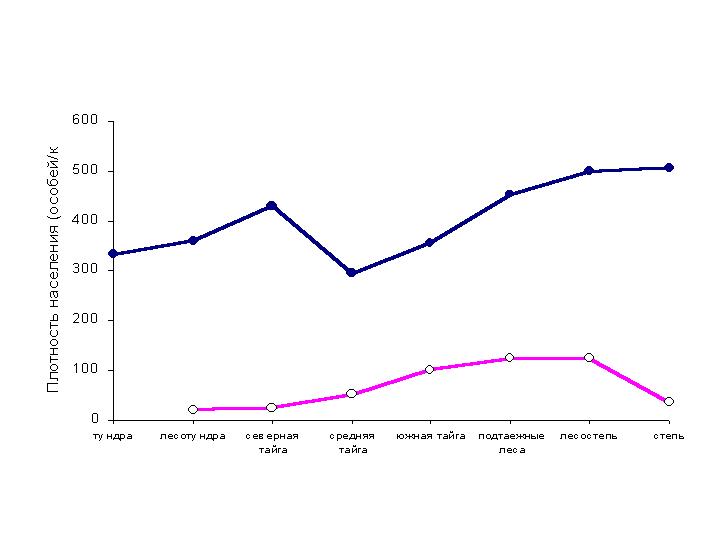
Рис. 1. Зональные изменения плотности населения птиц
Западно-Сибирской равнины.
Несомненно,
что все эти причины, обычно взаимосвязанные, приводят к увеличению летней
численности птиц в северном направлении. При этом заметное возрастание суммарного обилия
северотаежных птиц происходит в результате резкого увеличения численности
гипоарктических и бореально-гипоарктических видов, на фоне заметного снижения
численности многих бореальных видов.
В зимний период плотность населения птиц существенно
уменьшается в направлении от лесостепи и подтаежных лесов до северной тайги и
лесотундры. В степи обитает меньше птиц, чем в лесостепи, но больше, чем в лесотундре
и северной тайге. Видовое богатство населения птиц, выраженное количеством
встреченных видов, в летний и зимний сезоны изменяется сходным образом,
уменьшаясь от лесостепной до тундровой зоны (рис. 2). На фоне этой главной
тенденции прослеживается менее выраженное снижение видового богатства в
подтаежных лесах и степи. В подтаежных лесах это определяется малой площадью
подзоны, относительной монотонностью ее растительности и незначительной
представленностью хвойных лесов. Обеднение орнитокомплексов степной зоны
связано с выпадением из их состава некоторых лесных видов. При этом видовое
богатство северотаежных орнитокомплексов значительно больше, чем лесотундровых,
но несколько меньше, чем среднетаежных, как зимой, так и летом. Это
определяется значительным числом бореальных видов, распространенных в средней и
северной тайге, но не проникающих в лесотундру.
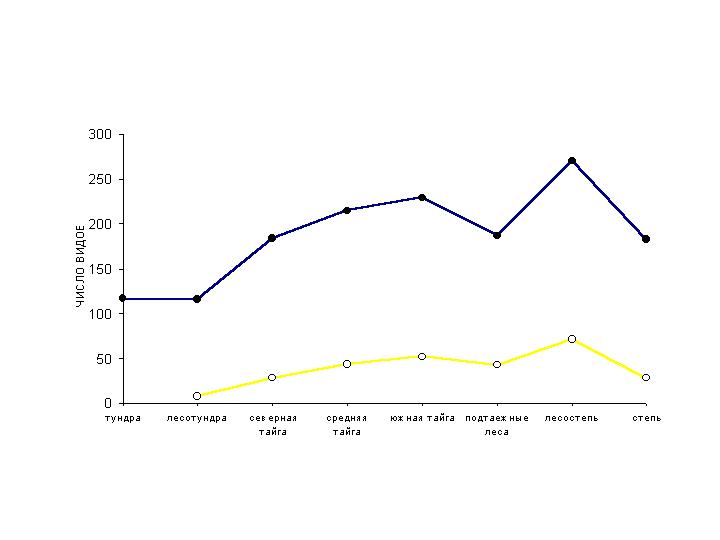
Рис. 2. Зональные изменения видового богатства населения птиц Западно-Сибирской
равнины.
Итак, обеднение летнего населения птиц в северном
направлении относительно невелико. Общее количество встреченных видов
уменьшается от лесостепной до тундровой зоны в 2,3 раза, а число особей в 1,5
раза. Зимой эта тенденция выражена значительно сильнее: от лесостепи до лесотундры
видовое богатство населения птиц сокращается в 10 раз, а их суммарное обилие –
в 6 раз. Столь резкое обеднение зимнего населения птиц с юга на север определяется
ухудшением защитных и кормовых свойств местообитаний, в том числе снижением
урожайности семян и ягод деревьев и кустарников (Данилов, 1952; Формозов,
1976). Кроме того, на Севере возможно воздействие низких зимних температур и
короткого светового дня.
Биомасса
населения птиц летом характеризуется наименьшими значениями в средней и южной
тайге. В этих подзонах доминанты по биомассе – чирок-свистунок (Anas crecca),
глухарь (Tetrao urogallus) и рябчик (Tetrastes bonasia). В
среднем по зонам и подзонам биомасса птиц увеличивается от средней и южной
тайги с продвижением к северу до тундры и особенно к югу до лесостепи (рис. 3).
Увеличение показателей в северной тайге, лесотундре и тундре, по сравнению со
средней и южной тайгой, определяется возрастанием численности водоплавающих
птиц и белой куропатки. Возрастание суммарной биомассы птиц в подтаежных лесах,
лесостепи и степи связано с большей антропогенной трансформацией ландшафтов и
увеличением численности врановых, особенно грача и серой вороны (Corvus
cornix). Зимой общая биомасса птиц наиболее высока в лесотундре, в связи с
миграциями белых куропаток из южной тундры, и в подтаежных лесах, где в значительном
количестве концентрируется тетерев (Lyrurus tetrix). От южной до северной тайги
доминируют рябчик, тетерев и глухарь, и только в северной тайге - белая
куропатка, но численность всех видов тетеревиных в таежных подзонах невелика.
Поэтому биомасса населения птиц в таежных подзонах гораздо меньше, чем в
лесотундре, подтаежных лесах и лесостепи. Степная зона характеризуется
минимальной суммарной биомассой птиц, здесь, как и в лесостепи, доминируют
только синантропные виды: сорока и сизый голубь (Columba livia).
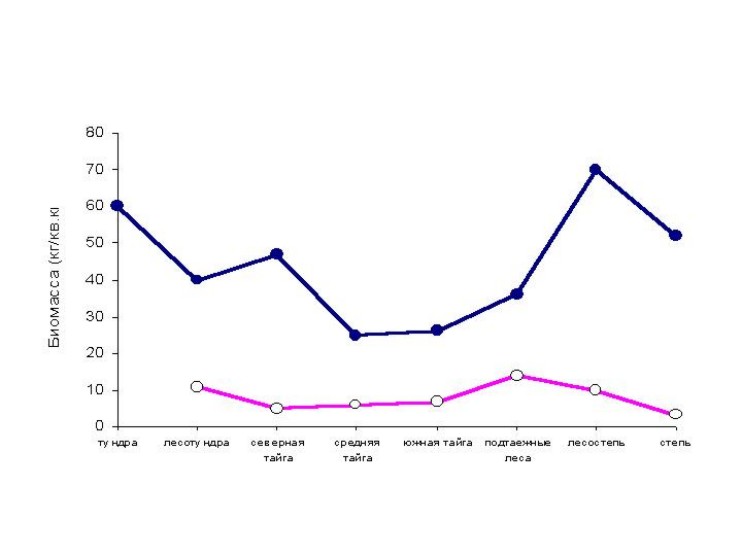
Рис. 3. Зональные изменения биомассы населения птиц
Западно-Сибирской равнины.
Фаунистический состав летних орнитокомплексов существенно
изменяется в связи со сменой зональных природных условий. Так, доля арктических
видов резко уменьшается в лесотундре и северной тайге, по сравнению с тундрой,
а еще южнее становится минимальной (рис. 4). Доля сибирских видов наиболее
высока в сообществах птиц лесотундры и северной тайги, лишь немногим меньше – в
средней и южной тайге, но резко уменьшается в тундре и подтаежных лесах, а
особенно – в лесостепи и степи. Участие европейских видов в орнитокомплексах
выше в южной тайге, подтаежных лесах и лесостепи, где больше распространены
мелколиственные леса, а к югу и особенно к северу оно резко уменьшается. Доля
транспалеарктов минимальна в южной и средней тайге, где большую площадь
занимают леса, особенно темнохвойные, но возрастает к северу и югу по мере
увеличения открытости ландшафтов. Только в тундровой зоне участие
транспалеарктов в населении птиц вновь уменьшается, здесь они уступают первое
место арктическим видам.
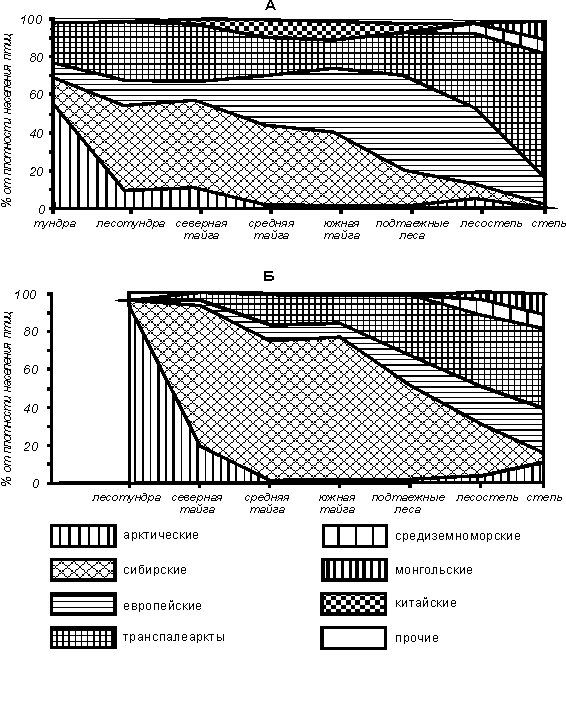
Рис. 4. Зональные изменения фаунистического состава населения птиц
Западно-Сибирской равнины.
В
результате такого распределения по градиенту зональности в летний период
представители отдельных типов фауны имеют не перекрывающиеся области преференции,
в каждой из которых одни из них преобладают над другими. Так, арктические виды
преобладают по численности в тундровой зоне, сибирские – от лесотундры до южной
тайги, европейские – в подтаежных лесах и лесостепи, а транспалеаркты – в
степной зоне. В зимний период состав орнитокомплексов более однородный, чем
летом, и на обширной территории от северной тайги до подтаежных лесов формируется
сибирский фаунистический состав населения птиц. В лесотундре зимой преобладают
арктические виды, а в лесостепи и особенно в степи – транспалеаркты при высокой
доле европейских видов.
Ярусное
распределение населения птиц в летний период характеризуется преобладанием
наземно-кормящихся птиц, доля которых к северу и югу от средней и южной тайги
возрастает (рис. 5). Следует учитывать, что птиц, добывающих корм на берегу
вблизи уреза воды, например трясогузок (род Motacilla) и многих куликов
(подотряд Сharadrii), мы также относим к группе птиц, кормящихся на земле.
Следующая по значимости группа – кронники. Их доля в орнитокомплексах
уменьшается к северу и югу от средней и южной тайги. В остальных ярусах
держится значительно меньше птиц, в порядке убывания их доли в населении это
группы кустарниковых, водных и кормящихся на стволах деревьев видов. Широтные
изменения их участия в орнитокомплексах соответствуют структуре растительности
и обводненности местообитаний. В зимний период всей лесной зоне (от северной
тайги до подтаежных лесов) свойственно преобладание кронников, которое сильнее
всего выражено в средней и южной тайге. В лесотундре, лесостепи и степи, как и
летом, доминируют птицы, кормящиеся на поверхности земли. Роль птиц, кормящихся
на стволах деревьев, в зимних орнитокомплексах по сравнению с летними заметно
возрастает, особенно в таежных подзонах, а кустарниковых птиц – уменьшается.
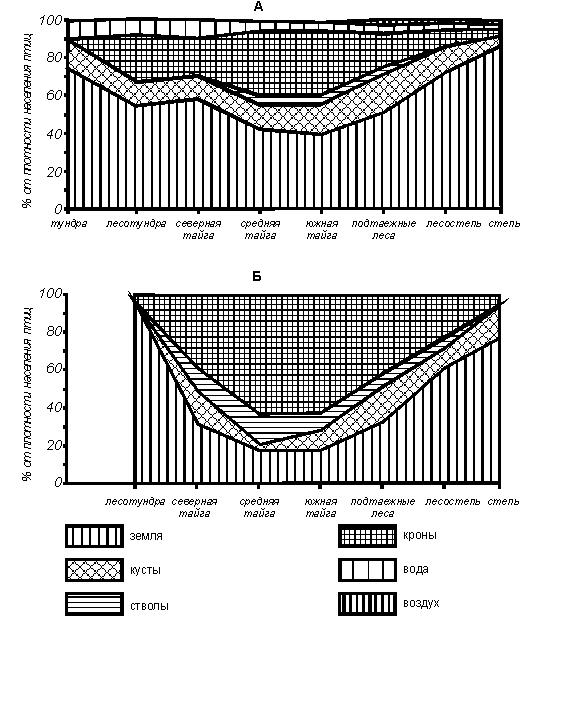
Рис. 5. Зональные изменения ярусного распределения населения птиц Западно-Сибирской
равнины.
Наибольшую
часть летнего рациона птиц составляют беспозвоночные (рис. 6). В южной тайге и
особенно к северу от нее в рационе птиц становятся все более значимыми
вегетативные части растений, поскольку возрастает участие в орнитокомплексах
представителей отрядов гусеобразных (Anseriformes) и курообразных
(Galliformes). Доля семян и плодов, повсеместно небольшая, увеличивается в
лесостепной и степной зонах, где больше синантропных птиц. Частично
потребляемые ими корма антропогенного происхождения включены в единую группу с
семенами и плодами. Позвоночные тоже мало значимы в летнем питании птиц, но их
роль несколько возрастает как в тундре, лесотундре и северной тайге в связи с
увеличением численности чаек, крачек и поморников, так и в лесостепи и степи,
что определяется возрастанием обилия врановых птиц, тяготеющих к агроландшафтам,
поселкам и городам.

Рис. 6. Зональные изменения состава кормов, потребляемых населением
птиц Западно-Сибирской равнины .
Зимой
рацион резко изменяется, в нем повсеместно преобладают семена и плоды. В южной
и средней тайге заметную долю в питании птиц составляют вегетативные части
растений, потребляемые лесными тетеревиными. В северной тайге и лесотундре их
роль особенно возрастает за счет более высокой численности белой куропатки по
сравнению с южнее расположенными территориями. Участие беспозвоночных в рационе
больше там, где выше обилие синиц, поползня и пищухи (в средней, южной тайге и
подтаежных лесах) и резко уменьшается к северу и югу. Доля позвоночных
становится заметной в подтаежных лесах, лесостепи и особенно в степи, т.е. там,
где больше синантропных врановых птиц.
ОБСУЖДЕНИЕ
В
летний период с продвижением с юга на север Западно-Сибирской равнины наиболее
заметные зональные изменения населения птиц наблюдаются в северной тайге.
Главные признаки, сближающие северотаежные орнитокомплексы с лесотундровыми и
даже с тундровыми, следующие: относительно высокие значения плотности населения
и биомассы; преобладание птиц, собирающих корм на земле и воде; заметное
значение вегетативных частей растений и позвоночных животных в рационе птиц;
высокая доля транспалеарктических и арктических видов. В более южных
орнитокомплексах эти показатели ниже. Лишь видовое богатство птиц наиболее
заметно сокращается при смене северной тайги лесотундрой. Последнее согласуется
с результатами исследований Чернова (1988, 1999), которые показали, что именно
в лесотундре происходит смена наиболее эволюционно продвинутых таксонов более
архаичными. По мнению того же автора, при снижении конкуренции со стороны
представителей наиболее прогрессивных отрядов, в первую очередь
воробьинообразных (Passeriformes), наиболее богатых видами в умеренных широтах,
представители эволюционно менее продвинутых отрядов, в первую очередь
ржанкообразных (Charadriiformes), ценотически замещают их в субарктических
наземных экосистемах, достигая здесь наибольшего биологического прогресса.
Сходная
тенденция прослеживается нами и в пределах отряда воробьинообразных при смене
среднетаежной подзоны северотаежной. Несмотря на то, что в северной тайге еще
сохраняются настоящие таежные местообитания, в теплое время года они заселяются
в основном гипоарктическими и бореально-гипоарктическими видами, более характерными
для кустарников и редколесий. Некоторые из этих видов, распространенные
полизонально, например, свиристель (Bombycilla garrulus), весничка (Phylloscopus
trochilus), овсянка-крошка, чечетка и др., обитают и значительно южнее
северной тайги. Однако в более низких широтах, будучи относительно менее
специализированными, они, возможно, испытывают конкурентное давление со стороны
более специализированных и многочисленных бореальных видов и не достигают
высокой численности. При ухудшении климатических условий с продвижением в более
высокие широты численность бореальных видов уменьшается, а гипоарктических и
бореально-гипоарктических, наоборот, возрастает. Последние, широко
распространенные в Субарктике, реализуют ряд своих адаптаций, которые связаны с
увеличением внутрисуточного периода кормодобывания, высокой трофической и
топической лабильностью и совмещением основных фаз годового цикла (Зимин, 1988;
Головатин, 1992; Рябицев, 1993). Кроме того, многие виды, обитающие в
Субарктике, в период гнездования способны смещаться в южном направлении при
особо неблагоприятных погодных условиях и сильном прессе хищников в тундровой
зоне (Данилов и др., 1984; Шутов и др., 1984) и задерживаться в северной тайге
(Вартапетов, 1984, 1998).
Следовательно,
в летний период на зональном градиенте от степей до тундр Западной Сибири
преобладает "блочный" принцип организации сообществ птиц. Он сводится
не столько к их постепенному обеднению или замещению одних видов другими, связанному
с зональной сменой природных условий, сколько к резкой смене бореальных
орнитокомплексов гипоарктическими при переходе от средней к северной тайге. При
этом северная граница древесной растительности при последовательном замещении
зональных северотаежных ландшафтов лесотундровыми и тундровыми не является существенным
рубежом в широтных изменениях населения птиц. Подобная смена бореальных
сообществ птиц гипоарктическими в Западной Сибири происходит южнее аналогичных
изменений растительности и ландшафтов в целом, а также сообществ мелких
млекопитающих, которые в северной тайге остаются преимущественно бореальными
(таежными). Птицы, как один из наиболее гибких компонентов экосистем, реагируют
больше на климатические, чем на ландшафтные изменения с продвижением в высокие
широты.
В
результате сообщества птиц северной тайги, лесотундры и тундры Западной Сибири
формируются в основном по правилу компенсации, выявленному ранее для тундровой
биоты (Чернов, 1985; Матвеева, 1998). Это правило определяется снижением
возможностей экологического викариата, напряженности конкурентных отношений и
увеличением относительной экологической емкости среды обитания. При продвижении
к северу количество видов, обитающих в определенных широтно-зональных условиях
уменьшается, но при этом численность некоторых из них возрастает. Т. е., уменьшение
видового богатства и разнообразия орнитокомплексов к северу от средней тайги
приводит к увеличению суммарного обилия птиц. На юге Западной Сибири летом при
переходе от лесной к лесостепной и далее к степной зоне не выявлено резких
изменений населения птиц. Это определяется высокой антропогенной,
преимущественно сельскохозяйственной освоенностью, которая возрастает в южном
направлении. Поэтому на зональном градиенте от южной тайги до степи происходит
постепенная смена лесных и опушечных видов птицами агроландшафтов.
Зимнее
население характеризуется постепенным обеднением состава и снижением
численности лесных видов в направлении от подтаежных лесов до лесотундры. В
лесостепи и степи, где зимние климатические условия несколько лучше, а давность
освоения и плотность населения человека значительно больше, преобладают
синантропные виды. Последнее определяется не только высокой концентрацией
местных врановых птиц в населенных пунктах, но и откочевкой синантропов из
расположенных севернее поселков и городов (Вартапетов, 1984; Блинов, 1998).
Итак,
в результате смены природных условий в зимний период по сравнению с летним на
Западно-Сибирской равнине с продвижением к северу особенно резко снижаются
видовое богатство и плотность населения птиц. Биомасса птиц в лесотундре,
наоборот, возрастает по сравнению с таежной зоной, за счет миграции белых
куропаток из тундровой зоны. В формировании зонального градиента зимнего
населения птиц доминирует принцип его обеднения от лесостепи до лесотундры за
счет снижения численности и постепенного исчезновения почти всех бореальных
видов. В летний период основной зональный тренд сводится к замещению бореальных
орнитокомплексов гипоарктическими в том же направлении. Полученные результаты
подтверждают предложенную ранее для умеренного климатического пояса Северной
Америки гипотезу о том, что пространственная структура зимних сообществ птиц
регулируется степенью суровости и изменчивости погодно-климатических условий, а
структура летних (гнездовых) сообществ отражает элементы межвидовой конкуренции
(Rotenberry et al. 1979). Кроме того, при переходе от летнего к зимнему
сезону для западносибирских орнитокомплексов в целом прослеживается увеличение
доли птиц, кормящихся на стволах и в кронах деревьев, преобладание растительных
и антропогенных кормов в рационе птиц, возрастание участия представителей
сибирского и арктического типов фауны.
БЛАГОДАРНОСТИ
Авторы
искренне благодарны вкладчикам Банка коллективного пользования по численности и
распределению животных ИСиЭЖ СО РАН В.А. Юдкину, В.С. Жукову, С.Г. Ливанову, Ю.А. Буйволову, С.А.
Соловьеву, К.В. Торопову, В.Н. Блинову, В.Г. Никитину, Т.Г. Джусупову, В.Н.
Плотникову, Е.Л. Шору, Ю.В. Бобкову, В.Г. Козину, Г.М. Тертицкому, И.В.
Покровской и К.В. Граждану, принимавшим участие в проведении зимних учетов
птиц.
Работа
выполнена при финансовой поддержке РФФИ (№ 99-04-50009).
СПИСОК ЛИТЕРАТУРЫ
Андреев А.В., 1980. Адаптация птиц к зимним
условиям Субарктики. - М.: Наука. 176.c.
Анзигитова Н.В., Бадаев С.А., Бурский О.В. и др., 1979. Некоторые зональные особенности населения птиц приенисейской северной тайги // Тез. докл. 8 Всес. зоогеогр. конф. М.: Наука. С. 92-93.
Базилевич Н.И., 1993. Биологическая
продуктивность экосистем северной Евразии. М.: Наука. 293 с.
Блинов В.Н., 1998. Врановые Западно-Сибирской
равнины. М.: KMK Scientific Press Ltd.
1-186 с.
Бутьев В.Т., 1970. Географическая изменчивость структуры населения птиц смешанных лесов Европейского центра СССР // Учен. зап. Моск. пед. ин-та им. В.И.Ленина. Т. 272. М. С. 57-71.
Вартапетов Л.Г., 1980. Внутриландшафтная структура
населения птиц северной тайги Приказымья // Проблемы зоогеографии и истории
фауны. Новосибирск: Наука. С. 156-166. - 1984. Птицы таежных междуречий
Западной Сибири. Новосибирск: Наука. 242.с. - 1998. Птицы северной тайги Западно-Сибирской
равнины. Новосибирск: Наука. 387 с.
Вартапетов Л.Г., Цыбулин С.М., Ливанов С.Г. и др., 2001. Классификация и пространственная организация зимнего населения птиц Западно-Сибирской равнины // Успехи совр. биол. Т.121, №6. С.615-625.
Гиляров М.С., Чернов Ю.И., 1974.
Некоторые итоги почвенно-зоологических исследований в СССР в плане Международной
биологической программы // Изв. АН СССР, серия биол. № 4. С. 563-579.
Гладков Н.А., 1958. О географической
изменчивости числа видов и численности особей // Проблемы зоогеографии суши.
Львов. С. 57-63.
Головатин М.Г., 1992. Трофические отношения воробьиных птиц на северной границе распространения лесов. Екатеринбург. 103 с.
Данилов Д.Н., 1952. Периодичность плодоношения и географическое размещение урожаев семян хвойных пород. М.-Л.: Гослесбумиздат. 142.с.
Данилов Н.Н., Рыжановский В.Н., Рябицев В.К., 1984.
Птицы Ямала. М.: Наука. 336 с.
Зимин В.Б., 1988. Экология воробьиных птиц
северо-запада СССР. Л.: Наука. 184 с.
Кищинский А.А., 1980. Понятие о гипоарктической и
эоарктической авифаунах // Экология, география и охрана птиц. Л. С. 121-133. -
1988. Орнитофауна северо-востока Азии. М.: Наука. 288 с.
Ливанов С.Г., Равкин Ю.С., 2001.
Мониторинг разнообразия наземных позвоночных государственного биосферного
заповедника «Катунский» (концепция, методы и вариант реализации // Труды
государственного природного биосферного заповедника «Катунский». Вып. 1.
Барнаул: Изд-во Алт. ун-та. С. 55-110.
Матвеева Н.В., 1998. Зональность в растительном покрове Арктики. Труды Ботанического ин-та. Вып. 21. С.-Петербург. 220 с.
Назаров А.А., 1968. Ресурсы и промысел боровой
дичи в северной части бассейна реки Таз // Ресурсы тетеревиных птиц в СССР. М.:
Наука. С. 52-53.
Паевский В.А., 1999. Адаптивная сущность
сезонных миграций: опасны ли для птиц их ежегодные перелеты? // Зоол. журн.
Т.78. Вып. 3. С. 303-310.
Пиминов В.Н., 1985. Перемещение белых куропаток
в период миграций. // Научные основы охраны природы Урала и проблемы
экологического мониторинга. Тез. докл. конф. молодых ученых. Свердловск. С. 45.
Портенко
Л.А., 1960. Птицы СССР. Ч. IV. М., Л.: Изд-во АН СССР. С. 1-415.
Птицы Советского
Союза. 1951-1954. Т. 1-6. М.: Наука.
Пузаченко Ю.Г., 1967. Географическая изменчивость
обилия и структуры населения птиц лесных биоценозов // Орнитология. Вып. 8. М.:
Изд-во МГУ. С. 109-122.
Равкин Ю.С., 1967. К методике учета птиц лесных ландшафтов // Природа очагов клещевого энцефалита на Алтае. Новосибирск: Наука. С. 66-75. - 1978. Птицы лесной зоны Приобья. Новосибирск: Наука. 288 с.
Равкин Ю.С., Вартапетов Л.Г., Юдкин В.А. и др., 1994. Пространственно-типологическая структура и организация летнего населения птиц Западно-Сибирской равнины // Сиб. экол. жур. N 4. С. 303-320.
Равкин Ю.С., Ливанов С.Г., Покровская И.В., 1999. Мониторинг разнообразия позвоночных на особо охраняемых природных территориях (информационно-методические материалы) // Организация научных исследований в заповедниках и национальных парках. М. С. !03-142.
Растительность Западно-Сибирской равнины., 1976.
Карта М. 1: 1500000 // Ред. И.С.Ильина. М.: ГУГК. 4 л.
Реймерс Н.Ф., 1990 Природопользование.
Словарь-справочник. М.: Мысль. -637.с.
Рябицев В.К., 1993. Территориальные отношения и
динамика сообществ птиц в Субарктике. Екатеринбург: Наука. 296 с.
Соколов Л.В., 1999. Популяционная динамика
воробьиных птиц // Зоол. журн. Т.78. Вып. 3. С. 311-324.
Степанян Л.С., 1990. Конспект орнитологической
фауны СССР. М.: Наука. 728 с.
Сыроечковский Е.Е., Рогачева Э.В., 1968.
Зональные особенности размещения и численности белой куропатки в приенисейской
северной тайге и лесотундре // Ресурсы тетеревиных птиц в СССР. М.: Наука. С.
77-78.
Формозов А.Н., 1976. Звери, птицы и их
взаимоотношения со средой обитания. М.: Наука. 309 с.
Цыбулин С.М., 1977. Географическая изменчивость населения птиц лесных ландшафтов зимой // Зоол. журн. Т. 56. Вып. 4. С. 588-594.
Чернов Ю.И., 1975. Природная зональность и
животный мир суши. М.: Мысль. 222 с. - 1985. Среда и сообщества тундровой зоны
// Сообщества Крайнего Севера и человек. М. С. 8-22. - 1988. Филогенетический
уровень и географическое распределение таксонов // Зоол. журн. Т. 67. Вып. 10.
С 1445-1458. - 1999. Класс птиц (Aves) в арктической фауне // Зоол. журн. Т.
78. № 3. С.276-292.
Штегман Б.К., 1938. Основы
орнитогеографического деления Палеарктики // Фауна СССР. Птицы. Т. 1 Вып 2.
М.-Л.: Изд-во АН СССР. 156 с.
Шутов С.В., Рябицев В.Н., Рыжановский В.Н., 1984.
Ежегодное перераспределение пеночки-веснички и пеночки-таловки в северной части
ареала // Вид и его продуктивность в ареале. Материалы 4-го Всес. совещ. Свердловск.
С. 89-90.
Jacobi A., 1919. Tiergeographie. Berlin-Leipzig. Vereinigung
wissenschaftlicher Verleger Walter de Vinter & C. 153 s.
Jarvinen
O., Sammalisto L., 1976.
Regional trends in the avifauna of Finnish peatland bogs // Ann. Zool. Fennici.
V. 13. P. 31-43.
Jarvinen
O., Vaisanen R.A., 1980.
Quantitative biogeography of Finnish land birds as compared with regionality of
other taxa // Ann. Zool. Fennici. V. 17. P. 67-85.
Kerr J.T.,
Packer L., 1997. Habitat heterogeneity
as a determinant of mammal species in high-energy regions // Nature. V. 385. P.
252-254.
Pianka E.R., 1966 Latitudinal gradients of species diversity: а review of concepts // American Naturalist. № 100. P.
33-46.
Rotenberry
J.T., 1978. Component of avian
diversity along a multifactorial climatic gradient // Ecology. V. 59. №4. P.
693-699.
Rotenberry
J.T., Fitzner R.F., Rickard W.H., 1979. Seasonal variation in avian community structure: differences in
mechanisms regulating diversity // Auk.
V. 96. № 3. P. 499-505.
Schmarda
L.K., 1853. Die geographische
Verbreitung der Tiere. Wien: Verlag Gerold und Sohn. 755 s.
РЕЗЮМЕ
CЕЗОННЫЕ ОСОБЕННОСТИ
ЗОНАЛЬНЫХ ИЗМЕНЕНИй
НАСЕЛЕНИЯ ПТИЦ
зАПАДНО-сИБИРСКОЙ РАВНИНЫ
Л.Г. Вартапетов, С.М. Цыбулин, С.П. Миловидов
Институт систематики и экологии животных СО РАН,
Новосибирск 630091
Зональные изменения летнего населения птиц Западно-Сибирской равнины от степной до тундровой зоны проявляются преимущественно в резкой смене бореальных орнитокомплексов гипоарктическими при переходе от средней к северной тайге. Обеднение населения, а также постепенное и последовательное замещение одних видов другими выражены в меньшей степени. Северотаежные, лесотундровые и даже тундровые летние орнитокомплексы отличаются от расположенных южнее более высокими показателями плотности населения и биомассы; большей долей птиц, собирающих корм на земле и воде; возрастанием доли вегетативных частей растений и позвоночных животных в рационе птиц; увеличением роли в сообществах транспалеарктов и арктических видов. Зимой от подтаежных лесов до лесотундры резко снижается суммарное обилие птиц и количество бореальных видов. На юге Западной Сибири (от южной тайги до степи) зональные изменения населения птиц менее выражены и более постепенны в связи с большей давностью и интенсивностью антропогенной трансформации местообитаний. Они сводятся в основном к смене лесных видов летом птицами агроценозов и зимой синантропами.
SEASONAL PECULIARITIES OF ZONE ALTERATIONS OF BIRD COMMUNITIES IN
WEST-SIBERIAN PLAIN
L.G. Vartapetov, S.M. Tsybulin, S.P. Milovidov
The zone alterations of summer bird
communities in West-Siberian plain from steppe zone up to tundra zone
pre-eminently appear in sharp replacement of Boreal ornithocomplexes by
Hipoarctic ones at the transit from middle to northern taiga. The
impoverishment of bird communities, and also the gradual and consequent
replacing of one species by others are expressed to a lesser degree. The
north-taiga, forest-tundra and even tundra summer ornithocomplexes differ from
located to the south ones by the more high indexes of density and biomass of
communities; the more fraction of birds foraging on ground and water; the
increase of the involvement of vegetative parts of plants and vertebrate
animals in a diet of birds; the increase of a role of Transpalearctic and
Arctic species in communities. In winter from subtaiga forests up to
forest-tundra the density of communities and the number of Boreal species
decreases. In the south of West Siberia (from a southern taiga up to steppe)
the zone alterations of bird communities are less expressed and more gradual,
it determines by the more remote and intensive anthropogenic transformation of
habitats. This alterations principally comes to the replacement of the wood
species by the birds of agricultural landscapes in summer and the synanthropic
species in winter.