УДК 591.9(5-013)+591.526+598.2/9
Классификация и пространственная организация зимнего населения птиц Западно-Сибирской равнины
Л.Г. Вартапетов, С.М. Цыбулин,
С.Г. Ливанов,
В.А. Юдкин, В.С. Жуков
Институт систематики и экологии
животных СО РАН, Новосибирск
На основе многолетних сведений по численности и распределению
птиц Западно-Сибирской равнины во всех типах ландшафтов и природных зонах, за
исключением тундровой, составлена классификация их зимнего населения. Выявлена
территориальная неоднородность орнитокомплексов, основные тренды которой
связаны с кормностью местообитаний и зональностью. К значимым факторам среды,
определяющим облик зимнего населения птиц, относятся также облесенность,
состав лесообразующих пород, укрытость,
провинциальность, антропогенное влияние, урожай семян и ягод деревьев и
кустарников, наличие тростникового яруса. Специфика
территориальной изменчивости зимних сообществ птиц по сравнению с летними
заключается в формировании инвазионного типа населения, в усилении воздействия
кормности местообитаний и ослаблении влияния зональности.
Введение
Формирование
зимнего населения птиц северной Евразии в значительной
степени определяется суровостью климатических условий и наличием
кормовых ресурсов. В зимний период облик населения птиц равнинной части лесной
зоны и гор юга Сибири становится довольно однородным и характеризуется
преобладанием таежных видов – представителей сибирского типа фауны [1, 5, 36].
Вместе с тем прослеживается дифференциация населения, связанная с широтно=зональными, высотно=поясными и долготно=провинциальными
различиями природных условий, а также с облесенностью,
кормностью, укрытостью и застроенностью местообитаний, и, кроме того, с
изменениями в составе основных древесных пород и развитии недревесных
ярусов растительности [7, 8, 21, 36-38].
Территориальная
неоднородность зимнего населения птиц лучше изучена в ландшафтно=зональном плане в Восточной и Северной
Европе [4, 5, 10, 13, 19, 50] и в горах юга Сибири, особенно Северо-Восточного
и Северного Алтая [21, 38]. Обширная территория Западно-Сибирской равнины до середины
1980-х гг. оставалась почти необследованной, за исключением Обь-Иртышской
лесостепи, городских и пригородных ландшафтов Новосибирска и Томска, а также
среднетаежного Зауралья [12, 15, 26, 32, 37]. Это определило необходимость проведения целенаправленных исследований зимнего населения птиц
Западно-Сибирской равнины. Хорошая изученность ландшафтно=зональной изменчивости летних
орнитокомплексов Западной Сибири, ее отдельных подзон или регионов [3, 6, 9,
22-25] позволила сопоставить пространственную структуру, классификацию и
организацию зимних и летних орнитокомплексов и проследить их сезонные изменения.
Районы и сроки работ, материалы и методы исследований
Птиц
учитывали на постоянных, но не строго фиксированных маршрутах, без ограничения
дальности обнаружения с последующим раздельно-групповым пересчетом на площадь
[20]. Работы проводили преимущественно в феврале с 1938 по
1942 г. и с 1975 по 1997 г. во всех природно-географических зонах Западной
Сибири, за исключением тундр. Всего обследовано 412 ландшафтных урочищ.
Общее число проанализированных вариантов населения, включая многолетние
сведения по Томску, Барнаулу и их окрестностям заметно больше - 670. Материал
собирали на 52 ключевых участках, места их расположения и годы обследования
указаны на рис. 1. В каждом местообитании за сезон длина учетных маршрутов
равнялась 10 км, а их общая протяженность - 7000 км. Часть
материалов собрана С.П. Миловидовым, Ю.А. Буйволовым, С.А. Соловьевым, К.В.
Тороповым, В.Н. Блиновым, В.Г. Никитиным, Т.Г. Джусуповым, В.Н. Плотниковым, Е.Л. Шором,
Ю.В. Бобковым, В.Г. Козиным, Г.М. Тертицким, И.В.
Покровской, К.В. Гражданом или заимствована из публикаций
[12, 18, 26, 30, 31].
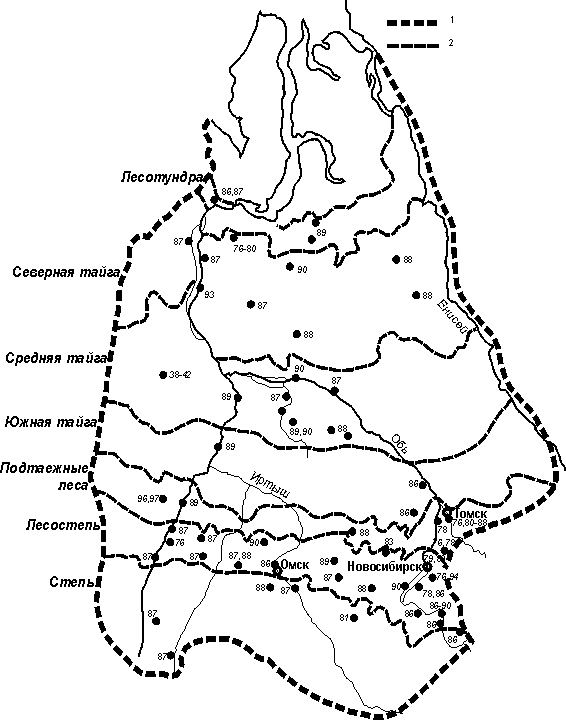
Рис.1. Места и годы проведения работ.
38-97 – 1938-1997 гг. Границы: 1 – Западно-Сибирской равнины, 2 –
природно-географических зон и подзон.
Для
классификации населения использовали один из методов кластерного анализа,
разработанный В.А. Трофимовым [33], согласно которому все рассматриваемые
варианты орнитокомплексов подразделяются на незаданное число классов, как по
наибольшему сходству вошедших в них вариантов населения, так и по степени их отличия
от остальных орнитокомплексов. В качестве меры общности
населения птиц использован коэффициент общности Жаккара,
модифицированный для количественных признаков [16, 49]. Более крупные классы
первого разделения всей совокупности вариантов населения птиц
иерархически разбивали на все более мелкие, до тех пор, пока для каждого из них
удавалось установить антропогенно=природный режим,
определяющий выделение того или иного класса [9, 25]. На основе такого
последовательного разделения составляли обобщенную классификационную схему населения
птиц.
Расчеты
проводили в три этапа. На первом из них классифицировали все 670 исходных
вариантов населения, т.е. многолетние данные не усреднялись. В некоторых
случаях результаты оказались плохо объяснимыми из-за совпадений
пространственных и временных различий орнитокомплексов. Неоднозначность и
разнонаправленность ряда классификационных объединений существенно затрудняли
интерпретацию полученных результатов. Вместе с тем на первом этапе выделены
инвазионные варианты, подтипы и инвазионный тип населения птиц. Их
выделение определялось значительным
сходством территориально разобщенных вариантов населения в годы с высоким
урожаем семян березы, ольхи, ягод рябины, боярышника и плодов сибирской яблони
за счет крайне высокого обилия инвазионных видов: чечетки, рябинника,
свиристеля и некоторых других. Сходство вариантов населения одних и тех же
местообитаний в годы с высоким и низким урожаем этих кормов оказалось гораздо
меньшим. Поэтому при всех последующих расчетах сохранялись инвазионные варианты
населения птиц.
На втором
этапе расчетов многолетние данные усредняли. Число анализируемых проб при этом
уменьшилось до 432, проведение расчетов стало легче, и главное, возросла
однозначность их интерпретации. Однако, классификация все
же имела некоторые недостатки, обусловленные малым сходством орнитокомплексов
при низкой численности птиц и случайностью встреч отдельных видов. В единые
классификационные подразделения часто входили варианты населения птиц резко
отличающихся ландшафтов. Интерпретация результатов второго этапа расчетов
оказалась излишне субъективной, так как классификационные подразделения
формировались на основе большого количества «волевых» решений (связанных с
перестановкой отдельных проб). Тем не менее, эти результаты послужили основой
для создания общей концептуальной схемы итоговой классификации.
На третьем
этапе расчетов пробы усреднялись в пределах выделов карты растительности
Западно-Сибирской равнины [27]. При этом случайно отличающиеся
орнитокомплексы объединялись с остальными в пределах одних и тех же
растительных формаций. Число анализируемых вариантов сократилось до 144. В
целом подтвердилась полученная на предыдущем этапе концептуальная
классификационная схема, а итоговая классификация оказалась лишенной недостатков,
выявленных ранее. Это связано с нивелировкой плохо объяснимых
годовых и территориальных различий в численности птиц, особенно связанных с их
малой заметностью и редкой встречаемостью. Остальные подходы и методы
исследований, в том числе выявления пространственной организации населения
птиц, неоднократно описывались в наших прежних публикациях [6, 9, 21, 23-25],
поэтому здесь не рассматриваются.
Классификация населения птиц
Процедура
классификации традиционно используется в биогеографических и экологических
исследованиях. Классификация орнитокомплексов может проводиться на заранее
выбранной, в том числе ландшафтной, основе путем последовательных усреднений их
характеристик. При этом предполагается полное совпадение территориальной
неоднородности ландшафтов или растительности и населения птиц, что чаще всего
оказывается неоправданным, поскольку территориальные изменения
орнитокомплексов, как правило, частично не совпадают с ландшафтно-зональными
условиями их местообитаний [6,9,22]. В отдельных случаях сообщества птиц
классифицируются по плотности населения, биомассе, видовому разнообразию или
иным обобщающим показателям, однако результаты бывают малоинформативными, поскольку
в одни группы могут попадать орнитокомплексы с резко отличающимся видовым составом
птиц и обитающие в существенно различающихся условиях. Несколько лучше подходит
для классификации населения состав доминантов, но и в этом случае высокий
субъективизм результатов препятствует их сравнимости. Наилучший результат дает
применение индексов сходства, которые давно используются при анализе общности
фаун в исторической биогеографии [14, 40, 42, 43, 47, 48, 51, 56, 57]. Наиболее
целесообразно применять показатели сходства, которые учитывают сходство и
различия населения не только на основе его видового состава, но и с учетом
обилия видов[39].
При
классификации орнитокомплексов по сходству типологический подход, используемый
в нашем исследовании, обеспечивает наибольшую степень упорядочения
представлений. Он позволяет выявить собственную дискретность
населения птиц, которая может определяться одним или несколькими факторами либо
их сочетаниями, территориальной сопряженностью, генезисом ландшафтов и многими
другими причинами. Схемы, полученные в результате классификации, весьма
информативны, не связаны с одним или несколькими заранее заданными признаками,
но требуют специальной интерпретации. Следует учитывать, что классификации, в
том числе экологические и биогеографические, скорее служат отправной точкой для
дальнейших исследований, чем описанием естественных структур. Основное назначение
подобных классификаций – выявить взаимосвязи между признаками фаун или
населения и факторами окружающей среды [35, 45].
Применяемая
нами автоматическая классификация, соотносит анализируемые объекты, в данном случае
– орнитокомплексы, по их сходству и различию. Она не связана заранее с выбором
признаков сообществ или факторов среды, пригодных для разделения анализируемой
выборки. Задачи такой классификации описываются Аркадьевым и Браверманом [2]. В самом общем виде они заключаются в
разделении больших совокупностей на несходные друг с другом группы, состоящие
из похожих объектов. Данный прием сжимает и упрощает анализируемую матрицу
признаков сообществ, выраженных коэффициентами их общности, позволяет выявить
преобладающие соотношения орнитокомплексов по их сходству и различию. При этом
анализ проводится в фиксированных границах, что обеспечивает определенность и
сравнимость получаемых представлений. Подобные задачи группировки сообществ
нередко решаются методами автоматической классификации в гидробиологических и
биогеографических исследованиях [17, 28, 34, 40, 41, 44, 46, 51, 52, 54, 56 и
др.].
Предлагаемая
ниже классификационная схема – идеализированный результат кластерного анализа,
как отражение полученных на его основе общих представлений о неоднородности
птичьего населения и определяющих ее антропогенно=природных
режимах. Для каждого классификационного подразделения приведены 5 видов,
преобладающих по обилию, с указанием доли каждого из них, плотность населения
птиц (особей/км2), число встреченных видов
и через косую - количество фоновых видов.
Система населения птиц
незастроенной суши
1. Л е с о т у н д р о в ы й т и п н а с
е л е н и я (лесотундры и лесов,
кроме темнохвойных северной тайги, а также лугово= и болотно=кустарниковых пойменных местообитаний от северной
до южной тайги; сероголовая гаичка 30%, белая
куропатка 24%, пухляк 14%, рябчик 6%, поползень 5%;
36, 26/7).
Подтипы населения:
1.1 - лесотундровых редколесий и
тундр (белая куропатка 98%, щур 0.8%, ворон 0.3%; 15, 3/1);
1.2 - северотаежных
не темнохвойных лесов (сероголовая гаичка 38%, пухляк 15%, белая куропатка 14%, рябчик 7%, поползень 6%;
45, 20/8).
Классы
населения: 1.2.1 - сосновых, лиственничных и
лиственнично-темнохвойных лесов (сероголовая гаичка 48%, рябчик 12 %, пухляк 11%, поползень и кедровка по 5%; 32, 18/8);
1.2.2
- мелколиственных и редкостойных
лиственнично-елово-березовых лесов и редколесий с
участками гарей и кустарниковых ивняков (сероголовая гаичка 30%, белая
куропатка 25%, пухляк 19%, поползень 6%, сорока 3%;
66, 18/10).
1.3. Подтип населения кустарниковых ивняков в поймах крупных рек от северной до южной тайги (белая куропатка 40%, чечетка
16%, снегирь 12%, пухляк 11%, сорока 6%; 23; 21/5).
2. Л е с н о й
т и п н а с
е л е н и я (лесов от средней тайги
до лесостепи, включая в северотаежные темнохвойные, преимущественно приречные
леса; пухляк 42%, большой пестрый дятел 8%, ополовник 7%, поползень и чечетка
по 6%; 91, 54/15).
Подтипы населения: 2.1 - северотаежных
темнохвойных, преимущественно приречных лесов (пухляк
39%, сероголовая гаичка 22%, кедровка 11%, чечетка 8%, поползень 6%; 103,
21/11);
2.2 -
темнохвойных и темнохвойно=сосновых
лесов средней и южной тайги (пухляк 49%, московка 11%, рябчик 7%, клест=еловик и поползень по 4%; 90, 30/14);
2.3 -
сосновых лесов от средней тайги до лесостепи (пухляк
44%, большой пестрый дятел 17%, поползень 8%, сорока 7%, ополовник 5%; 115,
45/12);
2.4 - внепойменных сосново= и темнохвойно=мелколиственных лесов
от средней тайги до лесостепи (пухляк 51%, московка 6%, ополовник и поползень
по 5%, большой пестрый дятел 4%; 73, 36/11);
2.5 -
мелколиственных и пойменных сосново=березовых
лесов от средней тайги до лесостепи (пухляк 34%, ополовник 13%, чечетка 12%,
поползень 7%, сорока 4%; 85, 46/17).
3. Л е с о п о л е в о й т и п н а с е л е н и я (полей, чередующихся с перелесками и
лесополосами, от южной тайги до степей, включая пойменные луга в сочетании с
ивняками в пределах подтаежных лесов и лесостепи, а также степные сосняки;
сорока и чечетка по 16%, пухляк 15%, ополовник и полевой
воробей по 9%; 34, 42/8).
Подтипы населения: 3.1 - полей, перелесков от
южной тайги до лесостепи и пойменных лугов, ивняков в подтаежных лесах и
лесостепи (чечетка и пухляк по 20%, ополовник, снегирь
и сорока - по 12%; 33, 39/7);
3.2 -
полей, лесополос в лесостепи и степи с проникновением к северу до средней тайги
по участкам припоселковых полей и лугов (полевой
воробей 27%, сорока 25%, пуночка 8%, чечетка 7%, князек
6%; 39, 30/10);
3.3 -
сосновых лесов в степной зоне (князек 39%, сорока 24%,
пухляк 19%, большой пестрый дятел 12%, большая синица 2%; 21, 10/4;).
4. Л у г о в о=с т е п н
о й т и п н а с е л е н и
я (степей и лугов в лесостепной и степной зонах
(чечетка 24%, пуночка 22%, сорока 15%, полевой воробей 12%, белокрылый
жаворонок 11%; 20, 26/5).
5. В е р х о в о б о л о т н ы й т и п н а с е л е н и
я (верховых, переходных,
бугристых и аапа=болот от северной до южной тайги,
включая южнотаежные внепойменные низинные болота;
пухляк 38%, большой пестрый дятел 14%, тетерев 8%, кедровка и сероголовая
гаичка по 6%; 6, 23/1).
Подтипы
населения: 5.1 - северотаежных комплексных бугристых, верховых и аапа=болот (сероголовая гаичка
38%, кедровка 18%, большой пестрый дятел и чечетка по 12%, тетерев 8%; 3,
10/0);
5.2 -
средне= и южнотаежных верховых, переходных и внепойменных низинных болот (пухляк
44%, большой пестрый дятел 15%, тетерев 7%, трехпалый дятел и чечетка по 5%; 8,
22/2).
6. Б о л о т н о=з а й м
и щ н ы й т и п н а с е л е н и я (заросших тростником озер и малых рек, займищ
и осоково-тростниковых болот от подтаежных лесов до степей; князек 92%, усатая
синица 4%, чечетка 1%, белая куропатка 0.8%, пуночка 0.7%; 34, 15/2).
7. И н в а з и о н н ы й т и п н а с е л е н и я (березово=хвойных
и осиново=березовых
лесов, ивняков, чередующихся с лугами и полей с перелесками, а также
диффузной городской застройки и парков от средней тайги до лесостепи при
высоком урожае семян березы, ольхи и ягод рябины, боярышника, а также плодов
сибирской яблони; рябинник 23%, свиристель 11%, большая синица и чечетка по
10%, сорока 9%; 1028, 63/34).
Подтипы населения: 7.1 - средне- и южнотаежных березово=темнохвойных,
подтаежных и лесостепных березово=сосновых лесов при
высоком урожае семян березы (пухляк 50%, чечетка 14%, большая синица и ополовник
по 7%, поползень 3%; 628, 48/23);
7.2 - осиново=березовых лесов, ивняков,
чередующихся с лугами и полей с перелесками и лесополосами от южной тайги до
лесостепи при высоком урожае семян березы, реже – ольхи а также ягод рябины
(чечетка 34%, снегирь 16%, сорока 10%, пухляк 6%, ополовник 5%; 463, 58/30);
7.3 -
парков, скверов, лесопарков, кладбищ и городской диффузной застройки в
подтаежных лесах и лесостепи при высоком урожае ягод рябины, боярышника и
плодов сибирской яблони (рябинник 33%, свиристель 15%, большая синица 13%,
сорока 9%, снегирь 5%; 1945, 36/25).
СИСТЕМА НАСЕЛЕНИЯ ПТИЦ
ЗАСТРОЕННОЙ СУШИ
8. С е в е р н ы й т и п н а с е л е н и я (городов и поселков лесотундры и северной
тайги, включая промышленные ландшафты от лесотундры до средней тайги; домовый
воробей 71%, сорока 11%, большая синица 5%, серая ворона 4%, полевой воробей
2%; 110, 18/8).
Подтипы населения: 8.1 - многоэтажной городской
застройки (сизый голубь 82%, домовый воробей 16%, ворон 1%, серая ворона 0.8%;
19, 4/2);
8.2 - малоэтажной
городской застройки, поселков и промышленных ландшафтов (домовый воробей 72%,
сорока 11%, большая синица 5%, серая ворона 4%, полевой воробей 2%; 121, 18/7).
9. Срединный тип населения (городов и поселков от средней
тайги до степи; домовый воробей 45%, сизый голубь 14%, полевой воробей 12%,
большая синица 8%, на юго востоке - черная ворона 5%; 2326, 57/19).
Подтипы населения: 9.1 - поселков от средней
тайги до степи (домовый воробей 49%, полевой воробей 22%, сизый голубь 9%,
сорока 8%, большая синица 7%; 1846, 39/17);
9.2 -
городов в средней тайге и малоэтажной городской застройки в южной тайге и
подтаежных лесах (домовый воробей 75%, большая синица 10%, сизый голубь 5%,
сорока 4%, полевой воробей 3%; 1295, 22/11);
9.3 -
сплошной многоэтажной городской застройки в южной тайге и подтаежных лесах и
городов в лесостепи (домовый воробей 57%, сизый голубь 16%, большая синица 10%,
полевой воробей 6%, свиристель 3%; 3063, 37/15);
9.4 -
городских парков, скверов, кладбищ и диффузной застройки городов от южной тайги
до лесостепи (домовый воробей 36%, большая синица 22%, полевой воробей и сорока
по 8%, снегирь 5%; 820, 49/22);
9.5 -
застроенных садово=огородных
участков от южной тайги до лесостепи (полевой воробей 38%, большая синица 14%,
снегирь 13%, сорока 12%, домовый воробей 6%; 440, 34/17);
9.6-
городских свалок в лесостепи (домовый воробей 23%, сизый голубь 22%, на юго=востоке – черная ворона 21%,
галка 13%, полевой воробей 11%; 22195, 11/10).
Согласно
классификационной схеме наиболее важные различия в зимнем населении птиц
связаны с наличием застроенных и незастроенных ландшафтов, что отражается в
выделении двух соответствующих им систем орнитокомплексов. Система населения
птиц застроенной суши, в свою очередь, подразделяется на два типа: северный и
срединный, близкие по видовому составу. Облик их населения в основном
характеризуется преобладанием синантропных видов, популяции которых оседлы либо
прикочевывают в населенные пункты на зимовку в связи с большим количеством в
них антропогенных кормов и убежищ от неблагоприятных климатических условий. Суммарное обилие птиц для северного типа в 20 раз меньше, чем для
срединного. Это объясняется малым размером, возрастом и небольшим количеством
городов и поселков, неразвитостью сельскохозяйственного производства и резким
возрастанием суровости всех климатических условий в лесотундре и северной тайге
по сравнению с южнее расположенными подзонами и зонами.
Система
населения птиц незастроенной суши отличается значительно большей
разнородностью, здесь выделено семь типов орнитокомплексов. Формирование
четырех из них: лесотундрового, лесного, лесополевого и лугово=степного
– в основном связано с зональными изменениями растительности. Остальные три
типа: верховоболотный, болотно=займищный и инвазионный – преимущественно интразональные.
При этом выделение первых двух определяется ухудшением кормовых и защитных
свойств местообитаний для многих зимующих видов по сравнению с лесными
ландшафтами. Вместе с тем наличие благоприятных условий для кормежки некоторых
видов в тростниковом и кустарниковом ярусах обеспечивает формирование болотно=займищного типа населения.
Последний, инвазионный тип, характеризуется существенным возрастанием кормности
в занимаемых им местообитаниях в годы с высоким урожаем семян и ягод деревьев.
Итак, выделение
типов населения определяется совместным воздействием облесенности, кормности, укрытости местообитаний, зональными различиями последних, а
также застроенностью, урожаем семян и ягод деревьев и кустарников, а также
наличием тростникового яруса. Дальнейшее разделение на подтипы и классы, наряду
с сочетаниями указанных факторов, связано с влиянием состава лесообразующих
пород, мезорельефа, характера застройки и наличием парков, лесопарков и свалок.
В зимний
период по сравнению с летним, за счет более широкого
распределения массовых видов на фоне снижения общего видового богатства,
территориальная дифференциация населения уменьшается. Так, для летнего
населения выделяется 83 таксона классификации [24], а для зимнего только 38.
Дробность и детализация классификационной схемы уменьшаются. Если летнее птичье
население разделяется на системы, надтиповые группировки,
типы, подтипы, классы, подклассы и роды, то зимнее – только на системы, типы,
подтипы и классы, т.е. количество уровней классификации
уменьшается с семи до четырех. Кроме того, зимой заметно расширяется зональный
спектр природных ландшафтов, занимаемых отдельными
типами населения. Так, в зимний период по сравнению с летним на 1-2 подзоны или
на 1 зону дальше к северу распространены лесной, лесополевой,
лугово=степной и верховоболотный типы населения, а к югу – лесотундровый.
Такие
изменения определяются массовыми кочевками ряда зимующих видов птиц и резким
обеднением птичьего населения открытых ландшафтов. Территориальная
дифференциация зимних орнитокомплексов по сравнению с летними
уменьшается также в связи замерзанием водоемов, наличием мощного снегового
покрова, опадением листвы и снижением роли беспозвоночных в рационе птиц. В
конечном счете пространственные различия зимнего
населения птиц определяются преимущественно защитными свойствами местообитаний
и распределением растительных кормов, что отмечалось ранее для средней тайги
Зауралья [26]. В результате информативность рассмотренной классификации
(выраженная величиной учтенной дисперсии коэффициентов сходства) несколько
меньше по сравнению с аналогичной схемой для летнего населения (соответственно
39 и 45%).
Пространственная структура и
организация населения
Пространственно=типологическая структура населения птиц рассматривается как общий характер территориальных изменений орнитокомплексов, который определяется их взаимосвязями. Для выявления такой структуры использовали наибольшие значения сходства между выделенными при классификации типами населения. На схеме (рис. 2) эта структура демонстрируется рядами выделенных групп орнитокомплексов. Каждый из рядов связывается с воздействием основного структурообразующего фактора среды, полученная схема ориентируется в факторном пространстве. Итак, для получения и объяснения пространственно=типологических структур используются приемы классификации и ординации. Первые сводятся к выделению и интерпретации дискретных групп сообществ. С помощью вторых определяются основные тренды территориальных изменений населения птиц и факторов среды и выделенные группы орнитокомплексов определенным образом ранжируются в факторном пространстве. Именно сочетание классификации и ординации часто дает наилучший результат в биогеографических исследованиях, поскольку оно соответствует двойственной – дискретной и континуальной – природе сообществ [45].
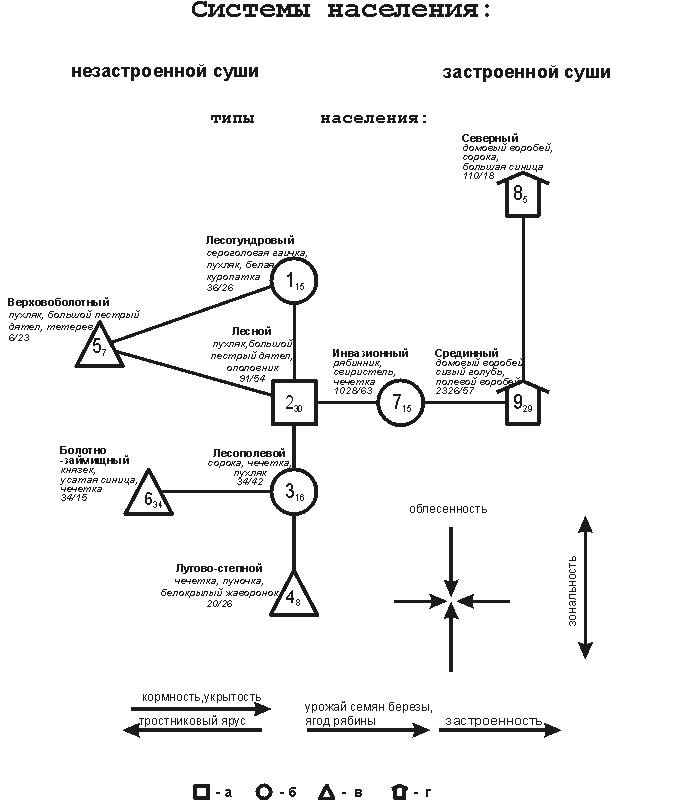
Рис. 2. Пространственно-типологическая структура зимнего населения
птиц Западно-Сибирской равнины.
Население птиц: а – лесных ландшафтов; б –
местообитаний, где чередуются участки лесов и открытых пространств; в –
преимущественно открытых ландшафтов; г – застроенных территорий. Цифрами
1-9 показаны номера типов населения. Индексы около этих цифр обозначают среднее
сходство проб, вошедших в данный тип. Рядом приводится название типа населения,
первые 3 преобладающие по обилию вида, плотность населения (особей/км2 и, через дробь – видовое богатство (количество
встреченных видов). Схема составлена в обратном масштабе: чем больше сходство
между типами, тем меньше расстояние между ними. Стрелки соответствуют основным
направлениям изменений орнитокомплексов и факторов среды.
На схеме
пространственно-типологической структуры населения птиц выделяются системы
орнитокомплексов: незастроенной суши (типы 1-7) и застроенной суши (типы 8, 9).
В первой из этих систем сверху вниз прослеживается зональная смена населения
птиц. При этом от первого к четвертому типу изменяется состав преобладающих
видов: обитатели кустарниковых тундр, лесотундровых редколесий и северотаежных редкостойных лесов (белая куропатка, сероголовая гаичка)
последовательно сменяются лесными видами (пухляк, большой пестрый дятел,
ополовник), лесополевыми (сорока и чечетка) и лугово=степными (пуночка и
белокрылый жаворонок). В системе населения птиц застроенной
суши в том же вертикальном направлении северный тип сменяется срединным
(соответственно с низким и высоким обилием преобладающих синантропных видов).
Изменения в
горизонтальном направлении в правой половине схемы определяются сменой
орнитокомплексов, связанной с увеличением кормности, укрытости
и застроенности ландшафтов в ряду: 2-й - 7-й - 9-й типы. При
этом лесной тип сменяется инвазионным, который занимает мелколиственные и
смешанные леса, участки диффузной городской застройки, лесопарки и парки от
средней тайги до лесостепи при высоком урожае в них семян березы и ольхи, ягод
рябины и боярышника, плодов сибирской яблони и увеличении численности
рябинника, свиристеля, чечетки и снегиря. Далее за этим типом следует срединный селитебный, занимающий преимущественно застроенные
местообитания, с преобладанием домового и полевого воробьев и сизого голубя.
В левой
части схемы тоже в горизонтальном направлении прослеживаются изменения
населения в связи со снижением облесенности, укрытости
и кормности при смене лесополевого типа населения болотно=займищным. Специфика
последнего – в преобладании князька и усатой синицы,
которые находят благоприятные кормовые условия в зарослях тростника и рогоза. Аналогичные изменения, связанные с резким ухудшением кормовых и защитных
качеств местообитаний, происходят при переходе от лесотундрового и лесного
типов к верховоболотному - орнитокомплексы последнего
формируются за счет крайнего обеднения населения первого и второго.
Итак,
пространственно-типологическая структура зимнего населения птиц
Западно-Сибирской равнины несколько упрощается по сравнению с таковой летнего.
Выделенные типы населения равномернее заполняют факторное пространство, в том
числе слабо выражена обособленность систем населения незастроенной и
застроенной суши. Для летних орнитокомплексов наиболее заметным является
зональный тренд изменений, а незначительные отклонения от него определяются
продуктивностью и кормностью ландшафтов. Для зимнего населения, наоборот,
сильнее выражены его территориальные изменения, связанные с кормностью и
укрытостью местообитаний, а зональные отличия выделяются слабее. Если в
формировании летнего населения доминирует правило независимого замещения [24],
то для зимнего наиболее значимым становится принцип обеднения - обогащения. Так, наибольшее суммарное обилие и видовое богатство птиц
характерны для инвазионного и срединного селитебного типов населения, а
максимальное видовое богатство – также и для лесного типа. Для остальных
типов населения с продвижением к северу, югу и в более открытые ландшафты эти
показатели резко уменьшаются.
Пространственная организация населения птиц рассматривается как связь территориальной неоднородности последнего с факторами среды. Эта связь оценивается величиной дисперсии коэффициентов сходства орнитокомплесов, учтенной (объясненной) каждым из выделенных факторов, их суммой, а также их неразложимыми сочетаниями или антропогенно-природными режимами [9, 24]. В результате может быть определена сила воздействия среды обитания на пространственные изменения орнитокомплексов, выявлена иерархия значимости этих факторов и установлена степень структурированности, упорядоченности сообществ. Под силой воздействия факторов среды на население птиц подразумевается не только причинно-следственная, но и любая корреляционная сила связи, определяющей здесь является степень зависимости изменений сходства орнитокомплексов и градаций рассматриваемых факторов.
Вместе с
этим, полученные оценки связей факторов среды с пространственной
неоднородностью населения птиц позволяют определить полноту объяснений
установленных закономерностей территориальных изменений орнитокомплексов.
Подобные исследования проводились и ранее, в том числе оценивались значения
факторов среды для формирования популяций и сообществ земноводных, птиц и
млекопитающих [11, 29, 53]. Позже сходные методы развивались и наиболее полно
использовались в наших работах [6, 9, 23, 24]. В данном
случае расчеты проводились по матрице 432х432 коэффициента сходства с
использованием случайных выборок в 100 проб с 8-кратной повторностью и с учетом
соотношения площадей выделов карты растительности Западно-Сибирской равнины.
При оценке силы связи факторов среды с территориальной
неоднородностью орнитокомплексов наиболее значимым оказалось влияние
облесенности и кормности (таблица). Несколько слабее воздействие состава
лесообразующих пород и укрытости местообитаний. Еще менее
значимо влияние зональности. Значительно более слабые факторы -
провинциальность и антропогенное воздействие (последнее выражается в виде
застроенности и распашки ландшафтов). Урожай семян и ягод деревьев и
кустарников, а также наличие тростникового яруса можно считать незначимыми
факторами в связи с малой площадью территорий, где они проявляются.
Итак,
степень воздействия отдельных факторов среды на пространственную неоднородность
населения птиц Западно-Сибирской равнины в зимний период изменяется по
сравнению с летним. Подобная оценка для летних орнитокомплексов была опубликована ранее [24]. Резко
возрастает воздействие кормности местообитаний (с 3 до 26%). Этот фактор зимой
сильно коррелирует с облесенностью и составом лесообразующих пород, т.е. лесные
ландшафты зимой всегда более продуктивны для птиц, чем открытые. Кроме того, в
лесах кормность возрастает с увеличением поверхности
питания, т.е. доли хвойных, особенно темнохвойных пород в составе древостоев,
что ранее показано для европейской части России и лесной зоны Приобья [8, 10]. Зональные различия орнитокомплексов зимой
несколько уменьшаются по сравнению с таковыми летом, что определяется не только
от-
Оценка силы и общности связи факторов среды с
территориальной
неоднородностью населения птиц Западно-Сибирской равнины
Фактор, режим |
Зимний период |
Летний период |
||
|
Средняя учтенная дисперсия, ± ошибка, % |
Интервал при 95%-ном уровне доверия |
Средняя учтенная дисперсия, ± ошибка, % |
Интервал при 95%-ном уровне доверия |
|
Облесенность |
28±0.8 |
25-30 |
27±1 |
24-29 |
|
Кормность, продуктивность |
26±1 |
23-29 |
3±0.3 |
2-4 |
|
Состав лесообразующих пород |
21±1 |
18-24 |
25±2 |
22-28 |
|
Укрытость |
21±1 |
19-24 |
—* |
— |
|
Зональность |
15±0.8 |
13-17 |
22±0.7 |
21-23 |
|
Провинциальность |
4±0.4 |
3-5 |
0.9±0.2 |
0.6-1 |
|
Антропогенное влияние В том числе: распашка застроенность |
3±0.4 2±0.3 2±0.5 |
2-4 1-2 0.5-3 |
3±0.4 3±0.4 0.7±0.2 |
2-4 2-3 0.2-1 |
|
Урожай семян и ягод деревьев
и кустарников |
1±0.2 |
0.5-2 |
— |
— |
|
Наличие тростникового яруса |
1±0.2 |
0.8-2 |
— |
— |
|
Мезорельеф |
1±0.1 |
0.7-1 |
5±0.6 |
3-6 |
|
Увлажнение, обводненность |
— |
— |
9±1 |
7-11 |
|
В с е г о факторы |
47±0.9 |
44-50 |
49±1 |
47-52 |
|
В с е г о режимы (по классификации) |
39±2 |
35-43 |
45±1 |
42-48 |
|
В с е г о
факторы и режимы |
55±1 |
52-58 |
59±1 |
56-62 |
личиями выборок (тундровая зона
зимой не обследована), но и большей широтой зонального распространения оседлых
и кочующих зимующих видов птиц, тогда как участие перелетных видов,
преобладающих в летнем населении, резко повышает его видовое и фаунистическое
разнообразие. При этом наиболее представительные по числу особей
типы фауны имеют различные области зональной преференции [24, 55], что
увеличивает широтную дифференциацию летнего населения птиц.
Заключение
Классификация
зимнего населения птиц Западно-Сибирской равнины выявляет его наиболее значимые
различия, которые определяют формирование двух систем орнитокомплексов:
застроенных и незастроенных территорий. На более низкой ступени классификации
выделено 9 типов населения в связи с воздействием облесенности, кормности, укрытости и зональной специфики местообитаний. Формирование
типов населения также определяется наличием тростникового яруса и урожаем семян
и ягод деревьев и кустарников. На еще более низких уровнях классификационной
схемы выделено 24 подтипа и 3 класса населения, что, наряду с сочетаниями выше
указанных факторов, определяется воздействием состава лесообразующих пород, мезорельефа,
характера застройки и наличия парков, лесопарков и свалок.
Пространственно=типологическая
структура орнитокомплексов демонстрирует 2 главных тренда территориальной
неоднородности населения птиц. Первый их них соответствует широтным изменениям
населения и определяется различиями в теплообеспеченности и зональной
спецификой ландшафтов. Второй тренд в основном связан с
кормностью местообитаний, которая, в свою очередь, определяется их
застроенностью и облесенностью. При оценке
силы и общности связи факторов среды с территориальной изменчивостью населения
птиц (с учетом площадей ландшафтов) наиболее значимым оказалось воздействие
облесенности, кормности, состава лесообразующих пород и укрытости
местообитаний. Несколько слабее выражены зональные отличия орнитокомплексов.
Существенно слабее влияние провинциальных и антропогенных различий ландшафтов,
а наиболее слабые и (или) локально проявляющиеся факторы – мезорельеф, урожай
семян и ягод деревьев и наличие тростникового яруса.
Территориальная
дифференциация зимнего населения птиц заметно уменьшается по сравнению с
таковой летом, что связано не только с отлетом большинства видов в более низкие
широты, но и с перераспределением зимующих птиц [8]. При этом
птицы не зависят от многих летних условий существования, в том числе
гнездования и кормодобывания, что определяется замерзанием водоемов, наличием
мощного снегового покрова, опаданием листвы и снижением роли позвоночных в
рационе [55]. Пространственные различия зимних орнитокомплексов, по
сравнению с летними, гораздо больше определяются кормовыми и защитными
свойствами местообитаний.
В
результате наиболее богатые типы птичьего населения формируются в лесных и
городских ландшафтах (за исключением лесотундровых и северотаежных), а также в
местах и в периоды высокого урожая семян и ягод деревьев и кустарников.
Последнее определяет обособление инвазионного типа населения птиц. Формирование
всех остальных типов населения преимущественно связано с резким снижением
суммарного обилия и видового богатства при переходе к слабо облесенным
и открытым ландшафтам. Таким образом, если летние орнитокомплексы в основном
формируются по правилу независимого замещения, то зимние – преимущественно по
принципу обеднения – обогащения [24].
Доля
растительных кормов (включая антропогенные) в зимнем
рационе птиц резко возрастает по сравнению с летним и становится преобладающей
[55]. При этом кормность местообитаний в зимний
период становится гораздо более значимой для формирования орнитокомплексов, а
влияние зональности, наоборот, несколько уменьшается по сравнению с летним
периодом.
Работа
выполнена при финансовой поддержке Российского фонда фундаментальных исследований.
СПИСОК ЛИТЕРАТУРЫ
1.
Андреев
А.В. Адаптация птиц к зимним условиям Субарктики. М.: Наука,
1980. 176 с.
2.
Аркадьев
А.Г., Браверман Э.М. Обучение
машины классификации объектов. М.: Наука, 1971. 192 с.
3.
Блинова Т.К.,
Блинов В.Н. Птицы Южного Зауралья: Лесостепь и степь. Т. 2. Новосибирск:
Наука, 1999. 288 с.
4.
Буйволов
Ю.А. // Изучение птиц СССР, их охрана и рациональное использование:
Тез. докл.
9 Всесоюз. орнитол. конф. Ч.1. Л., 1986. С. 102.
5.
Бутьев В.Т. // Уч. зап. Моск.
пед. ин-та им. В.И.Ленина.
Т. 272. М., 1970. С. 57.
6.
Вартапетов
Л.Г. Птицы таежных междуречий Западной Сибири. Новосибирск:
Наука, 1984. 242 с.
7.
Вартапетов
Л.Г. // Материалы Всесоюз. орнитол. конф. Ч.2. Кн.1. Минск,
1991. С. 102.
8.
Вартапетов
Л.Г. // Сиб. экол.
журн. 1995. Т.2. №2. С. 146.
9.
Вартапетов
Л.Г. Птицы северной тайги Западно-Сибирской равнины. Новосибирск:
Наука, 1998. 387 с.
10. Губин А.Г., Преображенская Е.С., Боголюбов А.С., Васюкова
О.В.. // Всесоюз. совещание по проблеме
кадастра и учета животного мира: Тез докл. Ч.3. Уфа,
1989. С. 69.
11. Дедов Б.Б., Дидык Н.К.,
Кузьмин И.Ф. // Научные основы охраны природы. Вып.
3. М., 1975. С.32.
12. Козлов Н.А. Птицы Новосибирска. Новосибирск:
Наука, 1988. 159 с.
13. Лазарева Н.С., Преображенская Е.С., Боголюбов А.С. //
Экология. 1988. №4. С. 34.
14. Ларина Н.И., Гурылева Г.М. //
Териология. М.: Наука, 1972, С. 186.
15. Миловидов С.П. // Вопросы биологии. Томск.: Изд-во ТГУ, 1978. С. 16.
16. Наумов Р.Л. Птицы в очагах клещевого
энцефалита Красноярского края. Автореф. дисс. ... канд. биол. наук. М.: МОПИ,
1964. 19 с.
17. Новые методы в тематической картографии
(математико-картографическое моделирование и автоматизация). М.: Изд-во МГУ,
1978. 128 с.
18. Пасхальный С.П. // Наземные позвоночные
естественных и антропогенных ландшафтов северного Приобья.
Свердловск, 1989. С. 18.
19. Преображенская Е.С., Бурский
О.В. // Материалы 6 Всесоюз. орнитол. конф. Кн.2. М.: Изд-во
МГУ, 1974. С. 290.
20. Равкин Ю.С. // Природа очагов клещевого
энцефалита на Алтае. Новосибирск, 1967. С. 66.
21. Равкин Ю.С. Птицы Северо-Восточного Алтая.
Новосибирск: Наука, 1973. 375 с.
22. Равкин Ю.С. Птицы лесной зоны Приобья. Новосибирск: Наука, 1978. 288 с.
23. Равкин Ю.С. Пространственная организация
населения птиц лесной зоны (Западная и Средняя Сибирь). Новосибирск: Наука,
1984. 262 с.
24. Равкин Ю.С., Вартапетов
Л.Г., Юдкин В.А., Миловидов С.П., Торопов К.В., Цыбулин С.М., Жуков В.С., Фомин Б.Н., Адам А.М., Покровская
И.В., Ананин А.А., Пантелеев П.А., Блинов В.Н.,
Соловьев А.С., Вахрушев А.А., Равкин Е.С., Блинова
Т.К., Шор Е.Л., Полушкин Д.М., Козленко А.Б., Ануфриев В.М., Тертицкий
Г.М., Колосова Е.Н. // Сиб. экол.
журн. 1994. № 4. С. 303.
25. Равкин Ю.С., Лукьянова И.В. География
позвоночных южной тайги Западной Сибири. Новосибирск: Наука, 1976. 338 с.
26. Раевский В.В. // Методы учета численности и
географического распределения наземных позвоночных. М.–Л., 1952. С. 294.
27. Растительность Западно-Сибирской равнины. Карта
М. 1: 1500000 // : Ильина И.С., Лапшина Е.И., Махно
В.Д., Романова Е.А.. М.: ГУГК, 1976. 4 л.
28. Сагайдачный А.Ю., Гиляров А.И., Матвеев В.Ф. // Журн.
общ. биол. 1977 Т. 38, № 2 С.
218.
29. Семенов Д.В., Леонтьева О.А., Павлинов И.Я. // Бюл. МОИП. Отд. биол. 2000. Т. 105. Вып.
2. С. 3.
30. Соколова А.А., Олейник А.М. // Фауна и
экология позвоночных Урала и их охрана. Челябинск, 1983. С. 49.
31. Стрельников Е.Г. Вопросы орнитологии: Тез. докл.
5=й конф. орнитологов Сибири. Барнаул, 1995. С. 173.
32. Торопов К.В., Фомин Б.Н., Козлов Н.А., Цыбулин С.М., Вартапетов Л.Г., Блинов В.Н., Жуков В.С.//
Пространственно=временная динамика животного населения. Новосибирск, 1985. С.
67.
33. Трофимов В.А. // Модели агрегирования
социально=экономической информации. Новосибирск, 1978 С. 91.
34. Уильямсон М. Анализ биологических популяций.
М.: Мир, 1975. 279 с.
35. Харвей Д. Научное объяснение в географии.
М.: Прогресс, 1974. 502 с.
36. Цыбулин С.М. // Зоол. журн. 1977. Т. 56. Вып.
4. С. 588.
37. Цыбулин С.М. Птицы диффузного города. Новосибирск:
Наука, 1985. 169 с.
38. Цыбулин С.М. Птицы Северного Алтая.
Новосибирск: Наука, 1999. 519 с.
39. Чернов Ю.И. // Зоол. журн. 1971. Т. 50. Вып.
7. С. 1079.
40. Шиленков В.Г., Воронов Г.А. // Вопросы экологии и
териологии. Пермь, 1973. С. 105.
41. Brett M.T., Wiackowski
K.G., Lubnow F.S., Mueller-Solger A., Elser J.J., Goldman
C.R. // Ecology. 1994. Vol. 75, №. 8. P. 2243.
42. Hagmeier E.M. // System. Zool. 1966. V. 15. №.4. P. 603.
43. Hagmeier E.M.,
Stults D.C.A. // Syst. Zool. 1964. V. 13 №3. P. 125.
44. Harris P.M. // Ecology. 1995. V. 76. №. 2. P. 544.
45. Hengeveld R. Dynamic biogeography.
46. Hinch S.G.,
47. Holloway
J.D., Jardine N. // Proc. Linnean. Soc. Lond.
1968. V. 179. №. 2. P. 153.
48. Jablonski B. // Ekol. polska.
1964. V. 10. №. 4. P. 307.
49. Jaccard P. // Bull.
Soc. Vaund. Sci. Nat. 1902.
Vol. 38. - P. 69.
50. Jarvinen O.,
Vaisanen R.A. // Ann. zool. fennici. 1980. V. 17. P. 67.
51. Kadmon R.H., Pulliam H.R. // Ecology. 1993. V. 74. N.
4. P. 977.
52. Kaiser
G.W., Lefkovitch L.P., Howden
H.F. // Canad.
J. Zool. 1972. V. 50. P. 1087.
53. Power D.M. // Ecology. 1975. V. 56. №. 3. P. 616.
54. Stouffer
P.C. Biergaard R.D. // Ecology. 1995. V. 75. №. 8. P. 2429.
55. Vartapetov
L.G. // Biodiversity and dynamics
of ecosystems in north
56. Wiens J.D. // Ecology. 1991. V. 72. №. 2. P. 479.
57. Zischer D.R. // System. Zool. 1968. V. 7. №. 4. P. 48.