ВВЕДЕНИЕ
В
орнитологии и орнитогеографии существуют проблемы, которые требуют предварительной
упорядоченности представлений о том, как формируются области распространения
птиц, от чего может зависеть общая численность их популяций, что и как
определяет их обилие на отдельных участках ареала. Список задач, для решения которых
важны ответы на эти вопросы, достаточно велик. В качестве только самых обычных
примеров можно привести оценку численности птиц при изучении ее многолетних
изменений или при характеристике распределения ресурсов того или иного вида,
выявление и уточнение ареалов, выполнение всевозможных прогнозов численности.
Решать эти задачи в той или иной мере необходимо при выполнении многих
прикладных работ. Наиболее распространенная из них – оценка воздействия
хозяйственных объектов на экосистемы.
Важность решения указанных задач не вызывает сомнений у
большинства исследователей. В то же время перечисленные и некоторые другие
подобные области знаний обладают двумя свойствами, существенно затрудняющими
решение. Первое из этих свойств заключается в чрезвычайной трудности оценок
того, насколько сформированные в них представления об изучаемом объекте
адекватны реальности. Если такие оценки и делаются, то, как правило, самими же
авторами этих представлений и на основе той же парадигмы, в рамках которой они
были получены. С другой стороны, исследователи, придерживающиеся различающихся
наборов исходных концепций и подходов, формируют и различающиеся представления
об одном и том же объекте (к примеру, различия в характеристике ресурсов
какого-либо вида). В подобной ситуации объективная проверка сделанных выводов
или моделей может быть выполнена лишь при неформальном их использовании в
реальной практической деятельности. Но вторым свойством этих областей знаний
как раз является их невостребованность обществом в настоящее время. Это значит,
что их невозможно проверить на практике, так как реально они в ней не
применяются. Ведь едва ли можно отрицать, что имеющиеся факты якобы практического
использования подобных разработок – это не что иное, как формальность, и само
использование не приводит ни к каким социальным или экономическим эффектам.
Пока же выводы или модели, полученные в упомянутых исследованиях, представляют
лишь познавательный интерес для узкого круга специалистов.
По
этим причинам в подобных исследованиях наиболее велика вероятность того, что
последовательное развитие взглядов в рамках одной парадигмы может стать простым
углублением заблуждений. Именно в таких изысканиях более,
чем где-либо требуется чрезвычайно тщательная предварительная методологическая
проработка и в первую очередь тщательный отбор исходных концепций и допущений.
Частью
такой проработки послужит формирование единой системы представлений о том, как
взаимодействие вида со средой реализуется в его пространственном размещении.
Эта система поможет оптимизировать упомянутые работы
как на стадии постановки задач, так и на стадии их решения. При постановке
задач можно легче выявить и отказаться от тех из них, которые в принципе не
имеют решения (к примеру, на карте относительно крупного масштаба обозначить
границу ареала вида, рассчитать уравнение регрессии обилия по фактору, который
не значим для вида, и т.п.). Тем самым будет исключено получение артефактов,
которые возможны в результате иллюзии решения нерешаемых задач. Кроме того, такая
система ограничит возможность принятия некорректных допущений, поможет определить
достаточный объем выборки и логичность планируемых сравнений.
К
настоящему времени для формирования системы представлений о механизмах пространственного
распределения птиц имеются широкие возможности. В литературе накоплено
достаточно много работ, посвященных дополнениям и уточнениям данных о географическом
распространении видов, неоднократно обсуждались экологические потребности
многих птиц, но самое главное – широко представлены и проанализированы обширные
количественные материалы по обилию птиц в разных условиях. Обобщение таких
сведений может дать весьма ценные знания.
В
данной работе анализируется лишь часть подобных сведений. Они касаются в основном
той стороны взаимодействия птиц со средой, которая приводит к неоднородности их
пространственного распределения. Этот аспект взаимодействий будет именоваться организацией
пространственного распределения животных. Под организацией пространственного
распределения вида (подвида) понимается набор имеющихся у него приспособлений
к среде и обусловленная этими приспособлениями реакция его на внешние факторы,
реализующаяся в определенном характере пространственной неоднородности обилия.
Для выявления такой организации необходимо решить по крайней мере три взаимосвязанные задачи. Прежде всего, важно сформировать представление о том, к существованию в каких условиях приспособлен вид. Для многих видов это можно сделать на основе факторного анализа данных по обилию в различных условиях. Последнее возможно лишь после решения второй, принципиальной задачи – рационально определить параметры как для характеристики этой приспособленности, так и для оценки проявлений среды, которым соответствуют адаптации вида. Указанные параметры должны быть формализованными, но в то же время их оценка должна быть сравнительно простой. Третья задача заключается в выявлении характера зависимости обилия птиц от значимых факторов среды.
Глава 1. ЭКОЛОГО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА ПТИЦ И ОЦЕНКА СРЕДЫ
1.1. Проблемы рассмотрения среды при характеристике распределения птиц
С проблемой поиска и отображения факторов среды, определяющих
территориальное распределение животных, сталкиваются почти все исследователи,
которые изучают экологию отдельных видов или неоднородность многовидовых
комплексов. Знания об этих факторах необходимы при описании пространственной неоднородности
обилия какого-либо вида. Без таких знаний невозможны обоснованные прогнозы и, в
частности, экстраполяции, которые неизбежны при расчетах ресурсов изучаемой
группы животных. Представления об определяющих распределение вида факторах
будут необходимы и в том случае, когда исследователь попытается
проанализировать многолетнюю динамику численности. Эти представления,
во-первых, позволят корректно оценить численность (что в большинстве случаев возможно сделать лишь по показателям обилия) и,
во-вторых, дадут возможность отделить внешние причины изменений от
внутрипопуляционных. Кроме того, при иллюстрации распределения вида в форме
описания или географической карты неизбежно приходится дифференцировать
пространство по уровням обилия вида на нем. Такая дифференциация – одно из
частных проявлений проблемы рассмотрения среды.
Упомянутая проблема еще далека от разрешения. Традиционный
подход к ее решению, основанный на использовании ландшафтной, геоботанической
или иной подобной типологии, а чаще гибрида из 2-3 таких типологий,
малоэффективен. Созданные на ее основе представления о неоднородности обилия
вида на обширной территории редко бывают сколько-нибудь адекватными реальности.
Об этом косвенно свидетельствуют результаты специальных исследований: существенная
корреляция обилия вида с какой-либо из таких типологических характеристик
обнаруживается весьма редко [Курлавичус, 1986; Kurlavicius, 1995; Monjeau
et al., 1998]. Отсутствие заметных связей в большинстве
случаев вполне закономерно, ведь такие специалисты, как ландшафтоведы или
геоботаники, чьи типологии заимствуются зоологами для характеристики
распределения животных, при выполнении этих самых типологий из множества
возможных параметров территориальных объектов выбирают лишь весьма ограниченный
их набор. Причем этот набор они формируют только сообразно своим предметным
целям, а отнюдь не для того, чтобы характеризовать пригодность территории для
обитания того или иного вида животных. Вполне естественно, что в выбранных
таким образом наборах параметров только случайно могут оказаться значимые для
некоторых животных факторы или их маркеры.
При решении подобных задач корректнее дифференцировать
пространство на основе специфических параметров среды, функционально связанных
с изучаемым видом. Одной из возможностей формирования такой системы может быть
рассмотрение взаимоотношений животных с пространством на основе Хатчинсоновской
концепции экологической ниши [Hutchinson, 1965].
При этом вряд ли может вызвать серьезные возражения предположение о том, что в
поведении животных превалирует рациональность, т.е. общая стратегия поведения
направлена на минимизацию затрат энергии. Если принять эту точку зрения на
отношение животных к пространству, то ее логическим продолжением будет
следующее утверждение. В каждый конкретный промежуток времени внешняя среда в
восприятии животного дифференцируется всего на две части: пригодную для его
обитания в данный момент и непригодную. Первая – это часть пространства, на
которой значимые для определенного вида факторы формируют его нишу, независимо
от того, каковы остальные проявления среды. (Дифференциация животными среды по
остальным факторам нерациональна, если эти факторы незначимы.) Вторая часть –
все остальное пространство, которое животным не может использоваться и соответственно
в его восприятии никак не должно дифференцироваться.
Такой принцип дифференциации пространства подтверждается
поведением птиц. Способность птиц к полету позволяет у них отчетливее, чем у
других животных, проследить использование разобщенных фрагментов территории с
пригодным для себя набором условий, но различающихся по многим другим факторам.
При этом для них характерно отчуждение от всего промежуточного пространства
[Иваницкий, 1998]. Исследователю, изучающему распределение вида, вполне
достаточно характеризовать любой участок лишь по представленности на нем ниши
данного вида. Здесь вполне очевидна бесполезность более сложной дифференциации
пространства на основе упомянутых ранее типологий или их гибридов.
Сформировать представления о значимых признаках среды, т.е. о
тех, которые характеризуют параметры ниши данного вида, можно следующим
образом. Во-первых, необходимо рассматривать каждый участок, на котором
определено обилие вида, не как представителя какого-либо подразделения
ландшафтной, геоботанической или какой-то иной классификации или типологии, а
как географический индивид. Иными словами, необходимо анализировать все
известные индивидуальные характеристики участка, не ограничиваясь только
классификационными или типологическими. Во-вторых, посредством ординации или
ранжирования обследованных участков по градиенту обилия вида на них из всего
набора известных характеристик территорий выявить те, которые направленно изменяются
в этом ряду. В-третьих, выявленные таким образом факторы следует рассматривать
с точки зрения известных в настоящее время экологических потребностей видов.
Такой подход к поиску значимых факторов не нов, но его реализация
для крупных систематических групп животных затрудняется разной полнотой знаний
об экологических потребностях отдельных видов. По-видимому, значительные
пробелы в этих знаниях будут сохраняться еще долго. В настоящее время
сформировать единую для класса птиц систему представлений о таких факторах поможет
классификация видов по сходству их реализованных экологических ниш. О последних можно судить по особенностям распределения каждого
вида. Естественно, что при этом, как и при любой классификации, неизбежны потери
многих деталей в характеристиках отдельных хорошо изученных видов. Но в то же
время все без исключения классифицированные виды, даже те, экологические потребности
которых слабо изучены, будут иметь сравнимые между собой эколого-географические
характеристики.
Выбору критериев и выполнению такой эколого-географической
классификации видов и соответствующей ей типологии условий среды посвящены два
следующих раздела. Эти классификация и типология должны служить: во-первых, для
формализации и унификации представлений об организующих распределение животных
факторах; во-вторых, концептуальной основой моделирования обилия по проявлениям
факторов. Моделирование может выполняться для решения задач как познавательных
(например, оценка полноты знаний), так и практических (прогноз, экстраполяция и
т.д.).
1.2. Принципы и параметры эколого-географической классификации видов и типологии факторов среды
Классификация видов и типология факторов среды выполнялись при соблюдении следующих четырех принципов.
1.Виды птиц классифицировались исключительно по сходству их
реализованных экологических ниш. Сама классификация и ее
трактовка не подгонялись ни под какие типологии и классификации из других
областей знаний.
2. Типологические представления об условиях обитания птиц формировались только по результатам интерпретации этой классификации. В предлагаемой работе сначала классифицировались виды а затем, согласно полученной классификации, выполнялась типология значимых для птиц совокупностей факторов.
3. Предварительно все возможные параметры экологической ниши объединялись в несколько групп таким образом, чтобы выделенные группы не перекрывались по своему составу, и параметры, вошедшие в одну группу, не зависели от таковых из других групп. В этом случае условные результирующие оси групп в факторном гиперпространстве ниши априорно можно считать ортогональными. Это упростит условные измерения и другие характеристики ограничиваемого ими гиперобъема.
Исходя
из опыта предшествующих исследований, целесообразно выделить
пять групп параметров ниши. Первая объединяет совокупность поверхностей или
сред, свойства которых соответствуют адаптациям вида эффективно передвигаться
при кормодобывании. Такая совокупность поверхностей или сред, которые являются
наиболее удобной опорой для птицы при ее передвижении во время добычи корма,
будет именоваться благоприятным субстратом. Характеру субстрата, как правило,
соответствуют анатомические и морфологические особенности животных,
обеспечивающие эффективность сбора корма на нем. Наличие и количество
доступного для использования птицами благоприятного субстрата – один из самых
значимых параметров среды, характеризующих пригодность определенной площади для
обитания вида. Таксоны классификации видов, которые сформированы по сходству
этой совокупности видовых характеристик, названы субстратными группами. В свою
очередь все разновидности субстратов, которые отвечают потребностям видов
определенной субстратной группы, объединены в соответствующий тип субстрата.
При
изучении сообществ птиц установлено, что их видовое
разнообразие зависит не столько от видового состава и разнообразия
растительности, сколько от сложности и характера ее архитектоники [MacArthur et al., 1966].
Значимость архитектурных особенностей фитоценоза подтверждалась впоследствии
многими исследователями. Для зоогеографических объяснений подобную пейзажную характеристику
целесообразно применять не только в отношении растительности, но и ко всей
остальной совокупности условий на участке обитания. Следовательно, параметры
ниши, составившие вторую группу, в совокупности можно охарактеризовать как
"архитектурные" особенности места обитания. К этим факторам у птиц
существуют главным образом морфологические (особенности окраски, относительная
длина маховых и рулевых перьев и т.д.) и поведенческие приспособления, повышающие
эффективность защиты птиц от врагов. Такая совокупность условий среды будет
именоваться топоархитектурой. Предпочитаемая определенным видом топоархитектура
– это совокупность, свойства и взаиморасположение предметов и элементов
местности, которые соответствуют адаптациям данного вида, направленным на
защиту особей и их гнезд от неблагоприятных факторов. Таксоны, сформированные
на этой основе, названы топоархитектурными группами птиц. Соответствующие им
разновидности топоархитектуры объединены в типы.
Третья
группа параметров ниши объединяет определенные широтно(высотно)-климатические
условия, в которых вид приспособлен размножаться. Это в основном продолжительность
периода с определенными температурами и снежным покровом, стабильность погоды,
продолжительность светового дня и т.д. К указанным факторам у птиц имеется ряд
соответствующих физиологических и поведенческих адаптаций (сроки и особенности
прохождения репродуктивных процессов, линьки, миграционного состояния и т.д.).
Результат их реализации – широтные или высотные особенности обилия вида в
период размножения. Таксоны классификации, сформированные по этой совокупности
параметров, названы широтно-климатическими группами. Во многих случаях
климатические условия, к размножению в которых приспособлен вид, на равнине
формируются в определенном интервале широт. Высотные особенности распределения
птиц добавляются к широтным лишь тогда, когда горные
страны располагаются на удалении от благоприятных широт в сторону экватора. В
таком случае на определенных высотах могут формироваться климатические условия,
сходные с равнинными в районах, расположенных ближе к
полюсам.
Четвертая
группа проявляется в долготной специфике ареалов. Вероятно, она представляет
собой результат взаиморасположения территорий, где есть все условия для размножения,
и территорий, где имеется возможность благополучной зимовки, включая исторически
сформировавшиеся особенности успешных взаимоотношений с другими зимующими
видами и видами-резидентами. На этой основе сформированы долготные группы
видов. Долготная характеристика распределения вида в данном случае является
лишь маркером пока еще мало изученного комплекса приспособлений вида,
реализуемых им на зимовках.
Классификация
по пятой совокупности, трофической, в данной работе не проводится, поскольку
оценить количество и доступность пригодных кормовых объектов на больших
территориях при современных методах практически невозможно. Следует отметить,
что доступность корма отчасти зависит от количества соответствующего субстрата,
так как эффективное передвижение дает возможность птице эффективнее добывать
корм.
4. Классификация видов по каждой из выделенных групп параметров ниши выполнялась отдельно, независимо от других групп. Также независимо друг от друга осуществлялись и типология в каждой из групп факторов среды, соответствующих этим параметрам ниши. Поскольку используемые совокупности параметров ниши подбирались таким образом, чтобы их оси можно было считать ортогональными, отдельные составляющие классификации будут равными по рангу, а полная эколого-географическая классификация будет иметь многомерный ретикулярный характер (рис. 1). Точно такой же вид имеет и полная типология всей совокупности значимых факторов среды.
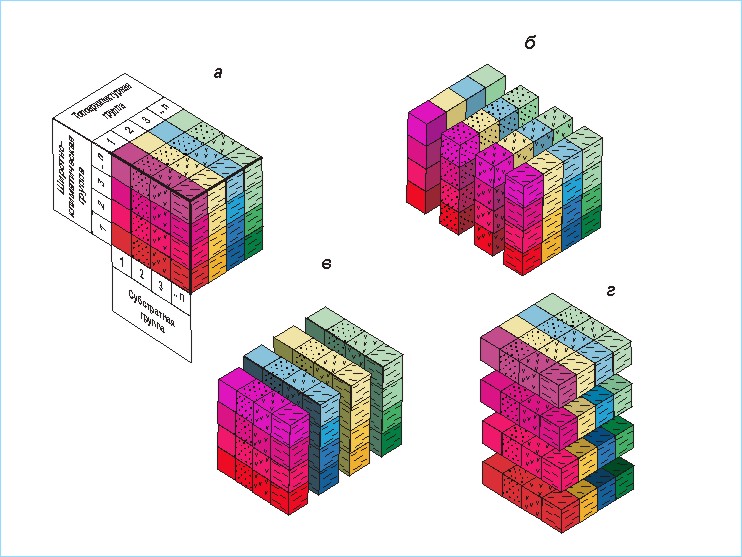
Рис. 1. Схема трехмерного
фрагмента многомерной ретикулярной классификации видов (а).
В
качестве примера отдельно показаны три составляющих ее классификации (раздвинуты: б –
субстратные группы; в – топоархитектурные
группы).
Такую классификацию А.А.Любищев [1982] считал наиболее удобной для решения подобных задач, называя ее комбинативной. По сравнению с традиционной иерархической ретикулярная (комбинативная) классификация имеет следующее преимущество. Как известно, распределение каждого вида уникально. При традиционной классификации стремление сохранить желательный объем информации порождает слишком большое число классов. Это чрезвычайно затрудняет восприятие такой классификации и ее осмысление. Если же агрегировать классификацию до такого количества таксонов, при котором она будет легко восприниматься, то потеря информации сделает результат банальным. Предлагаемая многомерная ретикулярная классификация позволяет легко воспринять многоклассовую конструкцию, поскольку каждая из составляющих классификаций может быть сравнительно простой. В то же время большое количество возможных комбинаций классов из каждой составляющей классификации дает возможность весьма полно отразить многообразие видовых ниш. В табл. 1 приведены комбинации для 186 видов. Тем самым даются примерные координаты их приспособленности в гиперпространстве факторов среды.
Т а б л и ц а 1
Эколого-географическая классификация птиц лесной зоны Западной
Сибири
|
|
Группа* |
|||
|
Вид |
Субстратная |
Топоархитектурная |
Широтно-клима-тическая |
Долготная |
|
Gavia arctica |
8 |
24 |
3 |
1 |
|
Podiceps nigricollis |
8 |
23 |
9 |
1 |
|
P. auritus |
8 |
23 |
8 |
1 |
|
P. cristatus |
8 |
23 |
9 |
1 |
|
Botaurus stellaris stellaris |
6 |
23 |
9 |
1 |
|
Ardea cinerea cinerea |
6 |
20 |
9 |
1 |
|
Anser anser |
6(7) |
23 |
9 |
1 |
|
Cygnus cygnus |
7 |
21 |
6 |
1 |
|
Anas platyrhynchos |
7 |
9 |
8 |
1 |
|
A. crecca |
7 |
4 |
4 |
1 |
|
A. strepera |
7 |
23 |
9 |
1 |
|
A. penelope |
7 |
4 |
6 |
1 |
|
A. acuta |
7 |
11 |
6 |
1 |
|
A. querquedula |
7 |
11 |
8 |
1 |
|
A. clypeata |
7 |
11 |
8 |
1 |
|
Aythya ferina |
8 |
23 |
9 |
1 |
|
A. fuligula |
8 |
11 |
7 |
1 |
|
A. marila |
8 |
24 |
2 |
1 |
|
Clangula hyemalis |
8 |
11 |
2 |
1 |
|
Bucephala clangula |
8 |
2 |
5 |
1 |
|
Melanitta nigra |
8 |
11 |
3 |
4 |
|
M. fusca |
8 |
11 |
3 |
3 |
|
Mergus albellus |
8 |
2 |
5 |
1 |
|
M. serrator |
8 |
24 |
5 |
1 |
|
Pandion haliaetus haliaetus |
1 |
20 |
7 |
1 |
|
Pernis apivorus |
1 |
4 |
8 |
3 |
|
Milvus migrans migrans |
1 |
17 |
7 |
2 |
|
M. m. lineatus |
1 |
17 |
7 |
5 |
|
Circus cyaneus cyaneus |
1 |
5 |
6 |
1 |
|
C. aeruginosus aeruginosus |
1 |
9 |
8 |
3 |
|
Accipiter gentilis buteoides |
1 |
4 |
5 |
4 |
|
Ac. g. schvedowi |
1 |
4 |
8 |
5 |
|
Buteo lagopus lagopus |
1 |
17 |
2 |
3 |
|
B. l. menzbieri |
1 |
17 |
2 |
6 |
|
B. buteo vulpinus |
1 |
5 |
7 |
4 |
|
|
1 |
5 |
7 |
1 |
|
Haliaeetus albicilla albicilla |
1 |
20 |
4 |
1 |
|
Falco peregrinus peregrinus |
1 |
17 |
7 |
4 |
|
F. p. calidus |
1 |
17 |
2 |
4 |
|
F. subbuteo subbuteo |
1 |
17 |
4 |
1 |
|
F. vespertinus |
1 |
5 |
7 |
4 |
|
F. tinnunculus tinnunculus |
1 |
5 |
8 |
4 |
|
Lagopus lagopus koreni |
5 |
9 |
3 |
5 |
|
L. l. maior |
5 |
9 |
9 |
4 |
|
Lyrurus tetrix tetrix |
5(3) |
5 |
5 |
5 |
|
L. t. viridanus |
5(3) |
5 |
7 |
4 |
|
Tetrao urogallus taczanowskii |
5(3) |
7 |
7 |
4 |
|
Tetrastes bonasia septentrionalis |
5(3) |
6 |
7 |
5 |
|
Coturnix coturnix coturnix |
5 |
11 |
9 |
3 |
|
Grus grus |
6 |
5 |
8 |
1 |
|
Porzana porzana |
6 |
9 |
8 |
3 |
|
P. parva |
6 |
23 |
8 |
4 |
|
Crex crex |
5 |
11 |
8 |
3 |
|
Gallinula chloropus chloropus |
7 |
9 |
9 |
1 |
|
Fulica atra atra |
7 |
23 |
9 |
1 |
|
Pluvialis squatarola |
5 |
12 |
1 |
1 |
|
P. apricaria |
5 |
12 |
3 |
4 |
|
Charadrius hiaticula |
5 |
14 |
3 |
1 |
|
Vanellus vanellus |
5 |
12 |
8 |
1 |
|
Tringa ochropus |
6 |
25 |
7 |
1 |
|
T. glareola |
6 |
16 |
3 |
1 |
|
T. nebularia |
6 |
5 |
6 |
1 |
|
T. totanus |
6 |
12 |
9 |
1 |
|
T. erythropus |
6 |
12 |
3 |
1 |
|
T. stagnatilis |
6 |
12 |
8 |
1 |
|
Actitis hipoleucos |
6 |
24 |
7 |
1 |
|
Xenus cinereus |
6 |
24 |
3 |
1 |
|
Phalaropus lobatus |
7 |
12 |
2 |
1 |
|
Philomachus pugnax |
6 |
12 |
3 |
1 |
|
Calidris temminckii |
6 |
21 |
3 |
1 |
|
Lymnocryptes minimus |
6 |
11 |
3 |
1 |
|
Gallinago gallinago |
6 |
11 |
6 |
1 |
|
G. megala |
6 |
4 |
7 |
6 |
|
G. media |
6 |
12 |
7 |
3 |
|
G. stenura |
6 |
11 |
5 |
6 |
|
Numenius arquata arquata |
6 |
4 |
8 |
2 |
|
N. ar. orientalis |
6 |
4 |
8 |
5 |
|
N. phaeopus phaeopus |
6 |
16 |
3 |
4 |
|
Limosa limosa limosa |
6 |
11 |
8 |
3 |
|
L. l. melanuroides |
6 |
11 |
8 |
6 |
|
L. lapponica lapponica |
6 |
12 |
2 |
3 |
|
L. l. menzbieri |
6 |
12 |
2 |
6 |
|
Larus minutus |
7 |
22 |
4 |
1 |
|
L. ridibundus |
7(5) |
23 |
8 |
1 |
|
L. heuglini antelius |
7(5) |
21 |
3 |
4 |
|
L. h. heuglini |
7(5) |
21 |
3 |
6 |
|
L. cachinnans cachinnans |
7(5) |
21 |
9 |
3 |
|
L. canus heinei |
7(5) |
21 |
4 |
1 |
|
Chlidonias |
1 |
23 |
9 |
3 |
|
Ch. leucopterus |
1 |
22 |
9 |
1 |
|
Sterna hirundo |
1 |
21 |
8 |
1 |
|
S. paradisaea |
1 |
21 |
2 |
1 |
|
Columba palumbus palumbus |
5 |
4 |
8 |
3 |
|
C. oenas oenas |
5 |
5 |
8 |
3 |
|
C. livia** |
5 |
19 |
9 |
1 |
|
Streptopelia orientalis |
5 |
4 |
9 |
6 |
|
Cuculus canorus canorus |
3 |
5 |
8 |
1 |
|
C. saturatus horsfieldi |
3 |
4 |
7 |
6 |
|
Asio flammeus |
1 |
11 |
4 |
1 |
|
Surnia ulula |
1 |
2 |
7 |
1 |
|
Strix nebulosa |
1 |
2 |
5 |
1 |
|
Apus apus apus |
1 |
17 |
8 |
3 |
|
Alcedo atthis |
8 |
18 |
9 |
1 |
|
Jynx torquilla |
3(5) |
5 |
7 |
1 |
|
Dryocopus martius |
4 |
1 |
7 |
1 |
|
Dendrocopos major |
4 |
1 |
7 |
1 |
|
D. leucotos uralensis |
4 |
2 |
8 |
4 |
|
D. minor minor |
4 |
4 |
7 |
3 |
|
D. minor kamtschatkensis |
4 |
4 |
7 |
6 |
|
Picoides tridactylus |
4 |
2 |
5 |
1 |
|
Riparia riparia |
1 |
18 |
4 |
1 |
|
Hirundo rustica |
1 |
19 |
9 |
4 |
|
Eremophila alpestris flava |
5 |
13 |
2 |
1 |
|
Alauda arvensis |
5 |
11 |
9 |
1 |
|
Anthus trivialis trivialis |
5 |
4 |
8 |
4 |
|
An. hodgsoni yunnanensis |
5 |
2 |
8 |
6 |
|
An. pratensis |
5 |
16 |
3 |
3 |
|
An. cervinus |
5 |
11 |
3 |
1 |
|
Motacilla flava thunbergi |
5 |
16 |
3 |
5 |
|
M. f. beema |
5 |
11 |
8 |
5 |
|
M. citreola citreola |
5 |
9 |
3 |
5 |
|
M. c. werae |
5 |
9 |
8 |
4 |
|
M. cinerea melanope |
5 |
25 |
7 |
6 |
|
M. alba |
5 |
15 |
4 |
1 |
|
Lanius collurio |
3 |
8 |
8 |
3 |
|
L. excubitor sibiricus |
3 |
5 |
5 |
6 |
|
L. ex. homeyeri |
3 |
5 |
8 |
3 |
|
Oriolus oriolus oriolus |
3 |
2 |
8 |
3 |
|
Sturnus vulgaris |
5 |
17 |
8 |
3 |
|
Perisoreus infaustus |
3 |
2 |
7 |
1 |
|
Garrulus glandarius glandarius |
3 |
2 |
8 |
3 |
|
G. g. brandtii |
3 |
2 |
8 |
6 |
|
Pica pica |
5 |
17 |
8 |
1 |
|
Nucifraga caryocatactes macrorhynchos |
3 |
1 |
7 |
6 |
|
Corvus monedula monedula |
5 |
17 |
8 |
3 |
|
C. frugilegys frugilegys |
5 |
5 |
9 |
3 |
|
C. cornix |
5 |
17 |
4 |
3 |
|
C. corax |
5 |
17 |
4 |
1 |
|
Bombycilla garrulus garrulus |
3(1) |
3 |
5 |
1 |
|
Locustella certhiola |
2 |
22 |
7 |
6 |
|
L. naevia naevia |
2 |
11 |
9 |
2 |
|
L. n. straminea |
2 |
11 |
9 |
5 |
|
L. lanceolata |
2 |
10 |
8 |
6 |
|
Acrocephalus schoenobaenus |
2 |
9 |
4 |
3 |
|
Ac. agricola brevipennis |
2 |
9 |
9 |
3 |
|
Ac. dumetorum |
2 |
8 |
8 |
5 |
|
Ac. arundinaceus arundinaceus |
2 |
23 |
9 |
3 |
|
Hippolais icterina |
3 |
2 |
8 |
3 |
|
H. caligata |
3 |
11 |
9 |
4 |
|
Sylvia borin |
3 |
4 |
8 |
3 |
|
S. communis |
3 |
8 |
8 |
4 |
|
S. curruca curruca |
3 |
6 |
7 |
4 |
|
Phylloscopus trochilus yakutensis |
3 |
10 |
5 |
3 |
|
Phylloscopus trochilus acredula |
3 |
10 |
8 |
3 |
|
Ph. collybita fulvescens |
3 |
4 |
6 |
5 |
|
Ph. borealis |
3 |
2 |
5 |
5 |
|
Ph. trochiloides viridanus |
3 |
1 |
8 |
4 |
|
Ph. trochiloides plumbeitarsus |
3 |
10 |
8 |
6 |
|
Ph. inornatus inornatus |
3 |
2 |
6 |
6 |
|
Ph. fuscatus fuscatus |
3 |
8 |
9 |
6 |
|
Regulus regulus regulus |
3 |
1 |
8 |
3 |
|
Ficedula hypoleuca |
3(1) |
4 |
7 |
3 |
|
F. parva parva |
1 |
1 |
7 |
4 |
|
F. p. albicilla |
1 |
1 |
7 |
6 |
|
Muscicapa striata striata |
1 |
2 |
8 |
2 |
|
M. s. neumanni |
1 |
2 |
8 |
4 |
|
Saxicola rubetra |
1 |
16 |
8 |
3 |
|
S. torquata maura |
1 |
16 |
7 |
4 |
|
Oenanthe oenanthe oenanthe |
5 |
15 |
4 |
1 |
|
Phoenicurus phoenicurus |
3 |
2 |
7 |
3 |
|
Luscinia luscinia |
5 |
8 |
8 |
3 |
|
L. calliope |
5 |
7 |
6 |
6 |
|
L. svecica svecica |
5 |
9 |
3 |
1 |
|
L. s. pallidogularis |
5 |
9 |
8 |
5 |
|
Tarsiger cyanurus cyanurus |
3 |
6 |
7 |
5 |
|
Turdus atrogularis |
5(3) |
7 |
5 |
4 |
|
T. pilaris subpilaris |
5(3) |
4 |
8 |
3 |
|
T. p. pilaris |
5(3) |
4 |
3 |
3 |
|
T. iliacus iliacus |
5(3) |
8 |
5 |
3 |
|
T. philomelos |
5(3) |
6 |
8 |
3 |
|
T. viscivorus viscivorus |
5(3) |
7 |
8 |
3 |
|
Aegithalos caudatus caudatus |
3 |
7 |
8 |
1 |
|
Parus montanus borealis |
3 |
1 |
7 |
1 |
|
P. cinctus |
3 |
2 |
5 |
1 |
|
P. ater ater |
3 |
1 |
8 |
1 |
|
P. cyanus hyperrhiphaeus |
3 |
8 |
9 |
4 |
|
P. major major |
3 |
4 |
9 |
1 |
|
Sitta europaea |
4 |
1 |
8 |
1 |
|
Passer domesticus domesticus |
5(3) |
19 |
8 |
1 |
|
P. montanus montanus |
5(3) |
17 |
8 |
1 |
|
Fringilla coelebs |
3(5) |
2 |
7 |
3 |
|
F. montifringilla |
5(3) |
2 |
5 |
1 |
|
Spinus spinus |
3 |
1 |
7 |
1 |
|
Acanthis flammea |
3(5) |
3 |
3 |
1*** |
|
Carpodacus erythrinus erythrinus |
3 |
8 |
8 |
1 |
|
Uragus sibiricus |
3 |
9 |
8 |
6 |
|
Pinicola enucleator |
3 |
2 |
5 |
1 |
|
Loxia curvirostra curvirostra |
3 |
1 |
7 |
1 |
|
L. leucoptera bifasciata |
3 |
2 |
5 |
1 |
|
Pyrrhula pyrrhula |
3 |
1 |
7 |
1 |
|
Coccothraustes coccothraustes |
3 |
2 |
8 |
1 |
|
Emberiza citronella citrinella |
5(3) |
5 |
8 |
3 |
|
Emberiza citronella erythrogenus |
5(3) |
5 |
8 |
5 |
|
Em. leucocephala |
5(3) |
3 |
7 |
6 |
|
Em. schoeniclus passerina |
3(5) |
9 |
3 |
5 |
|
Em. s. pallidior |
3(5) |
9 |
8 |
4 |
|
Em. s.incognita |
3(5) |
9 |
9 |
3 |
|
Em. pallasi polaris |
3(5) |
9 |
3 |
6 |
|
Em. rustica |
5(3) |
7 |
5 |
1 |
|
Em. pusilla |
5(3) |
3 |
3 |
5 |
|
Em. aureola |
5(3) |
16 |
7 |
6 |
|
Calcarius lapponicus |
5 |
12 |
2 |
1*** |
|
Plectrophenax nivalis nivalis |
5 |
17 |
1 |
1*** |
*Субстратные
группы птиц, адаптированных добывать корм при передвижении по субстратам следующих
типов: 1 - воздух; 2 - замкнутые цилиндрические вертикальные поверхности; 3 -
замкнутые цилиндрические горизонтальные поверхности; 4 - незамкнутые
вертикальные поверхности; 5 - незамкнутые горизонтальные твердые поверхности; 6
- незамкнутые горизонтальные рыхлые или мягкие
поверхности; 7 - поверхность воды; 8 - толща воды. В скобках приведен тип
субстрата, который дополнительно используется видом в гнездовой период или на
использование которого вид переходит в послегнездовое время.
Топоархитектурные группы
птиц, предпочитающих топоархитектуры следующих типов: 1 - сомкнутые рослые
древостои; 2 - редкостойные рослые древостои; 3 - редкостойные низкорослые
древостои и (или) редкостойные высокие кустарники; 4 - полесье; 5 – лесополье; приземный полог средней высоты среди: 6
- сомкнутых рослых древостоев, 7 - редкостойных рослых древостоев, 8 - редкостойных
низкорослых древостоев и (или) редкостойных высоких кустарников, 9) открытых
пространств; 10 - низкий приземный полог среди редкостойных
низкорослых древостоев и (или) редкостойных высоких кустарников; открытые пространства: 11
- с низким приземным пологом, 12 - без развитого приземного полога с редкими
куртинами низкорослой растительности и (или) редкими кочками, 13 - без
развитого приземного полога, 14 - лишенные растительности, 15 - без развитого
приземного полога с выделяющимися по высоте малыми элементами местности, 16 - с
низким приземным пологом с выделяющимися по высоте средними элементами местности,
17 - без развитого среднего приземного полога с выделяющимися по высоте
крупными элементами местности, 18 - с вертикальными обрывами; 19 - территории с
крупными антропогенными сооружениями; 20 – чередование обширных открытых
акваторий с малыми территориями, занятыми рослым древостоем; 21 - чередование
обширных открытых акваторий с малыми территориями, свободными от древостоя и
среднего приземного полога; 22 – акватории, сплошь занятые надводной
растительностью; 23 – чередование открытых акваторий и занятых надводной
растительностью, создающей надводный полог средней высоты; 24 - прибрежные
пресноводные акватории без растительности (вместе с литоралью или подтопленной
частью берега); 25 - берега водоемов и (или) водотоков среди полесья.
Широтно-климатические группы:
1 - эоарктическая; 2 - гемиарктическая; 3 - гипоарктическая; 4 -
трансумеренная; 5 - бореальная; 6 - северная умеренная; 7 - среднеумеренная; 8
- южная умеренная; 9 - особенности широтно-климатического предпочтения не
выявлены, так как основная часть ареала простирается южнее лесной зоны.
Долготные группы: 1 -
транспалеарктическая; 2 - крайнезападная; 3 - срединно-западная; 4 -
западно-срединная; 5 - восточно-срединная; 6 - срединно-восточная; 7 -
крайневосточная.
**
Полудомашняя форма.
***
Вид имеет голарктическое распространение.
В свою очередь, сравнительно простые типологии отдельных групп факторов воспринимаются так же легко. Комбинация из определенных представителей каждой такой типологии позволит легко дифференцировать пространство на пригодную и непригодную для конкретного вида части, т.е. отделить площадь, на которой представлена видовая ниша, от всей остальной площади. В результате любые территориальные выделы могут количественно характеризоваться на предмет их емкости для изучаемого вида. Этой характеристикой будет доля площади выдела, на которой представлена ниша вида.
Полные перечни полученных таксонов по каждой из классификаций и соответственно типологии факторов приведены в примечании к табл. 1. Номера видовых групп соответствуют номерам одноименных типов выделенных групп параметров среды. В данной работе предлагаемый подход продемонстрирован на примере эколого-географической классификации птиц, встреченных в лесной зоне Западной Сибири, так как эта территория наиболее обеспечена сравнимыми количественными данными по их обилию, а результаты анализа данных неоднократно публиковались [Вартапетов, 1984; Равкин, 1984; Малков, Равкин, 1985; Гуреев, 1985; Юдкин, 1986б]
1.3. Формирование таксонов классификации видов и типология факторов среды
Формирование всех групп предлагаемой классификации видов, а также создание представлений о типах проявления значимых факторов выполнено на основе обзора и обобщения публикаций, посвященных экологическим и географическим классификациям птиц. В классификационной схеме названия видов и подвидов даны по Л.С. Степаняну [1990]. В схеме вид дифференцирован на подвиды только в том случае, если при анализе выделяются более одной зоны преференции, которые совпадают с характеристикой распространения подвидов, изложенной в упомянутой публикации. В табл. 1 приведены виды, по численности и распределению которых имеется достаточный количественный материал.
1.3.1. Субстратные группы птиц и типы субстратов
Для формирования субстратных групп информация получена из характеристик экологических типов птиц, предложенных Л.П.Познаниным [1978]. В основном эти характеристики касаются способа передвижения при кормодобывании и отчасти места сбора корма. Различия в специализации передвижения птиц и нередкая ее сезонная изменчивость иногда создают некоторые трудности при решении, к какой группе отнести тот или иной вид. В настоящей работе определяющим послужил тот тип субстрата, на котором птица находится больше времени при кормодобывании и в первую очередь используемый птицей во время приближения ее к корму и в момент схватывания его. Для уточнения представлений о местах сбора корма использован ряд публикаций [Птицы Советского Союза, 1951-1954; Владышевский, 1980; Бурский, 1987; Преображенская, 1990; и др.].
Всех
птиц, добывающих пищу в полете, Л.П. Познанин разделил на
несколько типов в зависимости от его форм. Но поскольку все эти виды
приспособлены передвигаться при кормодобывании по воздуху, то независимо от
форм полета в настоящей работе они объединены в первую субстратную группу (см.
табл. 1). В нее включены только те виды, которые схватывают корм
преимущественно налету, не присаживаясь перед этим на иной субстрат.
Соответствующий данным видам тип субстрата – воздух в свободном пространстве (рис. 2).
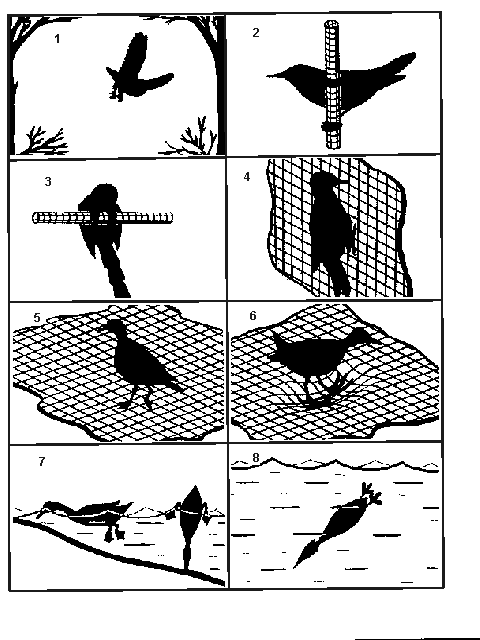
Рис.2. Схематическая
характеристика типов субстрата.
1 – свободное воздушное
пространство; 2 – замкнутые
цилиндрические вертикальные поверхности; 3
– замкнутые цилиндрические горизонтальные поверхности; 4 – незамкнутая вертикальная поверхность; 5 – незамкнутая горизонтальная твердая поверхность; 6 – незамкнутая горизонтальная
рыхлая и/или мягкая поверхность; 7
– поверхность воды; 8 – толща воды.
Л.П.
Познанин выделил совокупность птиц, добывающих пищу при помощи лазания по
болотным растениям (например, представители родов Locustella, Acrocephalus). Они приспособлены передвигаться не
только по болотным растениям, но и по жестким стеблям любых травянистых
растений и тонким ветвям кустарников, которые имеют ориентацию, близкую к вертикальной, и могут быть обхвачены пальцами птиц. На
основе этого нами выделена субстратная группа 2 – птиц, добывающих корм
преимущественно при передвижении по замкнутым
цилиндрическим поверхностям с вертикально ориентированной продольной осью
(диаметр поперечного сечения 5-20 мм). В свою очередь, все разновидности
субстратов с такими характеристиками объединены в одноименный тип. Общим свойством
этих поверхностей является то, что они могут быть обхвачены пальцами птиц и
только в таком случае могут служить для них опорой. Чем больше продольная ось
поверхностей отклоняется от вертикали, тем менее пригодны они для эффективного
передвижения птиц. Формально к данному типу субстрата следует относить все
подобные поверхности, если угол их продольной оси к горизонту превышает 45°.
Характеристики
большинства видов, отнесенных Л.П. Познаниным к обхватывающе-лазающим в ветвях
деревьев и кустов, послужили основой для формирования субстратной группы 3 – птиц,
приспособленных добывать корм при передвижении по замкнутым цилиндрическим поверхностям с горизонтально ориентированной
продольной осью (диаметр поперечного сечения 5 – 30 мм и более). Все
предметы с соответствующими характеристиками объединены в одноименный тип
субстрата. Так же, как и в предыдущем случае, данные поверхности могут быть
обхвачены пальцами птицы. Чем больше продольная ось таких поверхностей
отклоняется от горизонтали, тем менее пригодны они для птиц. Формально к этому
типу следует относить лишь те поверхности, продольная ось которых составляет с
горизонтом угол менее 45°.
Примерами
таких субстратов являются тонкие ветви деревьев и кустарников, а также жесткие
стебли трав с ориентацией в пространстве, близкой к
горизонтальной. Значительное разнообразие субстратов этого типа создается и
антропогенными предметами, особенно в урбанизированных ландшафтах. Сюда можно
отнести провода, проволочные ограждения, различные жерди. В качестве примера
можно привести также перила балконов в домах или бельевые веревки, натянутые на
балконах, которые существенно облегчают кормодобывание, в частности такого
вида, как Parus major. Интересным примером антропогенного субстрата может
служить большая куча беспорядочно перепутанной толстой проволоки, выброшенной на
свалку в промзоне г. Ноябрьска. Появление этого субстрата привлекло на кормежку
большое количество Phylloscopus trochilus, которые до того не имели
возможности там кормиться.
Характеристики
выделенной Л.П.Познаниным совокупности когтелазающих по стволам деревьев птиц послужили
основой для субстратной группы 4 – птиц, адаптированных добывать корм при
передвижении по вертикальным незамкнутым шероховатым поверхностям. К этому типу
субстрата можно отнести деревянные стены строений, участки обрывов или скал, а
также поверхности толстых стволов деревьев или столбов и любые другие
шероховатые поверхности с ориентацией, близкой к
вертикальной. Условно сюда можно отнести все подобные поверхности,
расположенные под углом более 45° к горизонтали.
При
оценке того, относить ту или иную поверхность к определенному типу субстрата,
необходимо учитывать, что незамкнутость ее в данном случае может быть весьма условной.
При этом рассматривать характеристики какой-либо обширной поверхности необходимо
в масштабе размеров изучаемой птицы. Так, если какая-либо поверхность и замкнута,
но диаметр ее поперечного сечения в несколько раз больше того, при котором она
может быть обхвачена пальцами птицы, то для передвижения птицы могут
использовать эту поверхность только как незамкнутую. А поскольку такие
поверхности обладают теми же свойствами, что и незамкнутые, то их следует относить
к данному типу субстрата. В качестве примера можно привести поверхность
толстого ствола дерева, по которой такие виды, как Dendrocopos spp., Sitta spp. и т.п., передвигаются точно так же, как и по любой незамкнутой
поверхности.
Для
всех бегающих, ходящих и прыгающих по земле птиц благоприятными для
передвижения являются не только поверхность земли, но и другие субстраты,
которые можно охарактеризовать как относительно ровную горизонтальную
поверхность (например, плотно слежавшаяся или плавающая на воде растительность,
поверхность плотного мохового или лишайникового покрова, плоские крыши строений
и т.п.). Однако способ обнаружения корма внутри этой группы птиц различается.
Одни виды обнаруживают его только с помощью зрения (к примеру, Motacillidae). Другие виды, имея удлиненный
чувствительный клюв, еще и зондируют верхние слои размягченного субстрата
(многие Charadrii), как правило,
приуроченного к переувлажненным участкам, рыхлому моховому покрову или плавающему
на воде слою растительности. В последнем случае для движения существуют дополнительные
приспособления (удлиненные пальцы или цевка и т.д.). По этому признаку все виды
с подобными адаптациями объединены в следующие субстратные группы: 5 – птиц,
собирающих корм при передвижении по твердым
горизонтальным незамкнутым поверхностям; 6 – птиц, адаптированных собирать
корм при передвижении по рыхлым или
мягким горизонтальным незамкнутым поверхностям. Поверхности, обладающие
указанными характеристиками и ориентированные к горизонтали под углом менее 45°, формально следует относить
соответственно к одному из этих двух типов. Необходимо отметить, что наиболее
мелкие воробьиные птицы из-за малой массы своего тела одинаково легко передвигаются
по обоим типам горизонтальных поверхностей.
Л.П.Познанин
не дифференцировал водных птиц на ныряющих и неныряющих, хотя они передвигаются
при кормодобывании на различных глубинах водоемов и водотоков. Это впервые
учтено Д.Н.Кашкаровым [1944] при выделении им жизненных форм. Часть этих птиц
собирает корм преимущественно во время плавания по поверхности воды и в верхнем
ее слое на глубине, которую птица может достичь, опустив часть корпуса в воду и
оставаясь на плаву. Ныряющие виды при сборе корма передвигаются в толще воды на
глубине, значительно превышающей длину тела птицы. Данные виды в предлагаемой
классификации объединены в две группы (7 и 8). Соответствующие типы субстратов
– поверхность воды и толща воды. К первому типу формально
следует относить лишь ту поверхность воды, глубина под которой не превышает 1м.
Для наиболее крупных птиц пригодной можно считать поверхность, под которой
глубина составляет 0 – 1,5 м. Ко второму типу субстрата следует относить толщу
воды с глубиной 0,5 – 4,0 м.
1.3.2. Топоархитектурные группы птиц и типы топоархитектуры
Эти таксоны формировались в результате сравнительного анализа семи опубликованных классификаций видов по сходству их пространственного размещения в I половине лета, т.е. преимущественно в гнездовой период [Вартапетов, 1984; Равкин, 1984; Малков, Равкин, 1985; Гуреев, 1985; Юдкин, 1986б; Сметанин, 1989; Касыбеков, 1990]. Указанные работы выполнены на основе обширного количественного материала, собранного в разнообразных условиях и в разных географических точках. Они хорошо сравнимы между собой, так как материал получен по единой методике, а при классификационных расчетах использован единый алгоритм [Трофимов, Равкин, 1980].
В
каждой из публикаций приведены группировки видов со сходным характером неоднородности
обилия в соответствующей выборке (совокупности обследованных территориальных
выделов). При этом принималось во внимание, что констатация сходства распределения
каких-либо видов, имеющая место лишь в одной из публикаций, могла основываться
и на случайном стечении обстоятельств. Окончательное решение об объединении
видов в одну группу принималось в том случае, если сходство их распределения
или связь с определенным фактором демонстрировались неоднократно на разных выборках.
Авторы
анализируемых работ называют предпочитаемыми те выделы, в которых виды
определенной группировки встречены с наибольшим для выборки обилием. Нами для
каждого вида проведены сравнения условий в предпочитаемых выделах, причем
сравнивались только их индивидуальные топоархитектурные характеристики. На
основе этих сравнений сформированы представления о типах топоархитектуры. Таким
образом, виды, предпочитающие в разных районах площади со сходными
физиономическими характеристиками, объединены в топоархитектурные группы. В
свою очередь, физиономические характеристики послужили основой для формирования
представлений о типе топоархитектуры.
Так,
в каждой из перечисленных работ выделены совокупности видов, предпочитающих те
или иные лесные территории. Но эти совокупности в каждой из классификаций
неоднородны и образуют несколько таксонов. При абстрагировании от
типологических ландшафтных трактовок авторов можно заметить, что в каждой из
классификаций присутствуют группы видов, предпочитающих различные леса с
заметным участием или преобладанием сомкнутого рослого древостоя. Кроме того,
во всех публикациях представлены совокупности, объединяющие виды по
предпочтению всевозможных разреженных лесов, а также других ландшафтных урочищ
с наибольшим для выборки участием редкостойной древесной растительности.
Неоднородность
в распределении видов последней группы сводится, главным образом, к различиям в
предпочтении, с одной стороны, разреженного рослого древостоя, а с другой –
разреженного низкорослого. Причем во втором случае виды встречаются со сходным
обилием, как в редколесьях, так и в разреженных высоких кустарниках. Указанные виды объединены в три группы, предпочитающие
топоархитектуру типов: 1 – сомкнутый
рослый древостой (кроны соседних деревьев соприкасаются, образуя сравнительно
плотный полог, высота деревьев более 18 м); 2 – редкостойный рослый древостой (кроны большинства соседних деревьев
не соприкасаются); 3 – редкостойный
низкорослый древостой (высота 1.5 – 18 м)
или редкостойный высокий кустарник (выше 1,5 м, рис. 3).
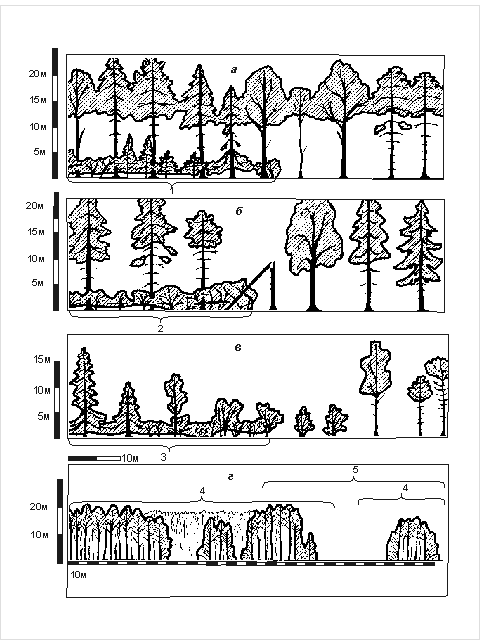
Рис. 3. Физиономические
профили топоархитектуры, сформированной
преимущественно древостоями.
а – сомкнутый рослый
древостой; а1
– то же со средним приземным пологом; б
– редкостойный рослый древостой; б2 –
то же со средним приземным пологом; в
– редкостойный низкорослый древостой и/или высокий кустарник; в3 – то же со средним приземным пологом;
г4 – полесье; г5 – лесополье; отдельные элементы этих топоархитектур по своим
характеристикам соответствуют также типам: г6
– сомкнутые древостои; г7 – редкостойные древостои; г8 – открытые пространства.
В
характеристики первых двух типов топоархитектуры помимо высоты и сомкнутости
деревьев следует включать еще наличие экранирующих элементов. Многочисленные наблюдения показывают, что почти все виды первых
двух топоархитектурных групп поселяются среди рослых древостоев только в том
случае, если с выбранного ими участка минимум на две стороны света не
просматривается открытое пространство. Выбранный птицей участок может
быть отделен от открытого пространства не только массивом деревьев,
расположенных по соседству с участком обитания, но и любыми другими высокими элементами
местности: скалой, обрывом, а в селитебном ландшафте – стенами высотных зданий.
Топоархитектура
первых двух типов широко представлена не только в лесах
и лесопарках, но и на болотах (по сухим облесенным гривам, на границе с суходолом,
кое-где на облесенной части болот), в пойменных уремах, в различных посадках,
скверах, а также в озелененных частях жилой или промышленной застройки. Топоархитектура
третьего типа представлена еще шире. В лесных
ландшафтах ее можно обнаружить на заболоченных участках, зарастающих вырубках,
у лесных опушек. Она преобладает по площади в низкорослых разреженных лесах, в
редколесьях и криволесьях, присутствует в зарослях кустарников и стлаников, на
различных болотах, в речных поймах, кустарниковых тундрах, посадках, садах и
т.п.
Во
всех анализируемых публикациях выделена группа видов, предпочитающих, по мнению
одних авторов, поля, чередующиеся с перелесками, а по
мнению других – луга с колками, луга по вырубкам или лесостепь. В отдельных
работах некоторые из этих птиц включены в группу предпочитающих пойменные леса
с луговинами. Общим для всех перечисленных урочищ является чередование открытых
пространств с участками леса, редколесья или массивов высоких кустарников.
Экологические особенности большинства этих видов таковы, что для укрытия гнезд,
охраны территории или при опасности они используют участки лесной растительности,
а как кормовую территорию – открытые пространства или опушки. Часть из этих
видов в большей степени приурочена к лесам с луговинами или полянами и в
меньшей степени к полям или лугам с перелесками. В последнем урочище они встречаются
или в перелесках, или в непосредственной близости от них. Другие, наоборот, в
большей степени приурочены к открытым участкам.
Проведенное
сравнение позволило объединить такие виды в две группы птиц, предпочитающих
соответствующие типы топоархитектуры: 4 – полесье;
5 – лесополье. Под полесьем
понимается площадь, занятая преимущественно древостоем с вкраплением в него
небольших по площади открытых участков (100 – 3000 м2).
Причем к этой категории относится лишь та часть древостоя, которая находится не
далее 100 м от границы с открытым участком. Лесопольем считается аналогичное по
масштабу сочетание сравнительно обширных открытых пространств с малыми
участками, занятыми древесной растительностью или высокими кустарниками (3000 –
10000 м2). У границы большого лесного
массива и обширного открытого пространства к полесью следует отнести приграничную
полосу леса на глубину до 100 м и сопредельную безлесную полосу на глубину до
20 м. Лесополье в этой местности будет занимать более широкую безлесную полосу
и 30 – 50-метровую лесную. Для многих хищных птиц средней и особенно крупной
величины, которые используют древостои лишь как место для устройства гнезда, а
корм добывают в открытых пространствах, к лесополью следует относить и ту часть
древостоя, которая удалена от опушки на более значительные расстояния.
Для
Обь-Иртышской долины выделена совокупность птиц, предпочитающих низинные
закустаренные болота [Юдкин, 1986б]. Для Южного Забайкалья многие из тех же видов
составили долинно-кустарниковый тип преференции [Сметанин, 1989], а для
Иссык-Кульской котловины они охарактеризованы, как предпочитающие караганники и
закустаренные степи [Касыбеков, 1990]. Во всех трех случаях на предпочитаемых
территориях одинаково проявляется один общий фактор: значительная площадь там
занята густыми зарослями кустарников. Большинство птиц встречается с высоким
обилием также в тростниковых займищах, зарослях крапивы или другого высокотравья.
Успешное обитание всех этих видов возможно лишь под пологом, создаваемым
растительностью на высоте 0,5 – 1,5 м над поверхностью земли или воды (рис. 4).
В дальнейшем полог, сформированный на такой высоте, будет именоваться пологом
средней высоты или средним приземным. Его могут создавать не только кустарники
или высокотравье, но и высокие кочки или низко расположенные ветви деревьев
(особенно хвойных).
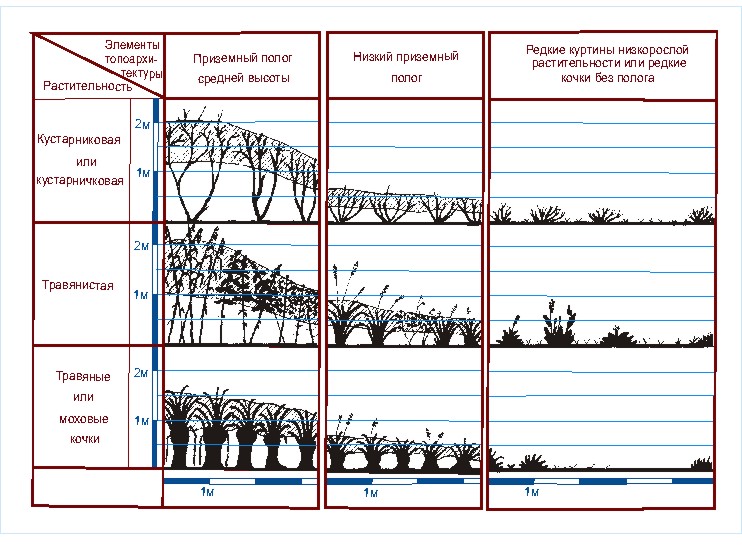
Рис. 4. Физиономические
профили топоархитектуры и ее элементов, образованных преимущественно кустарниками,
кустарничками, травами или мхами.
Разные
виды, приспособленные обитать под таким пологом, заселяют его в зависимости от
того, где он расположен. Анализ различий в распределении этих видов позволил
выделить четыре топоархитектурные группы птиц, предпочитающих полог средней высоты среди: 6 – сомкнутого рослого древостоя; 7 – редкостойного рослого древостоя; 8 – редкостойного низкорослого древостоя или
редкостойного высокого кустарника; 9 – открытых
пространств. Физиономические профили топоархитектуры этих типов отражены на
рис. 3 (а1, а2, а3).
Следует отметить, что многие из этих видов активно используют пространство не
только под этим пологом, но и над ним. Поэтому в набор параметров,
характеризующих топоархитектуру типов 6 – 9, наряду с характеристиками полога
обязательно входят и свойства соответствующих древостоев, кустарников или
безлесных пространств.
Для
успешного обитания многих видов необходим более низкий полог. Такой полог,
образованный на высоте 0,2 – 0,5 м над поверхностью земли или воды, будет
именоваться низким приземным пологом (см. рис. 4). Так же, как и средний полог,
его могут образовывать травы, кочки, кустарники, но более низкие. Низкий полог
часто формируется кустарничками. Помимо растительности его могут создавать и
антропогенные предметы. В качестве примера можно привести куски маскировочной
сетки, оставленные на редких кочках и куртинах низкой растительности после
военных учений.
Если
для многих видов, тяготеющих к древостоям или кустарникам, приземные условия
или безразличны, или им необходима относительно свободная поверхность почвы, то
для таких птиц, как Locustella lanceolata и некоторые Phylloscopus, непременным условием для обитания является наличие
низкого приземного полога (для указанных видов обычно на высоте 0,4 – 0,5 м).
Данные виды объединены в группу 10 – птиц, предпочитающих редкостойные низкорослый древостой и/или высокий кустарник с низким
приземным пологом. Необходимо уточнить, что к этому типу топоархитектуры
следует относить древостои и кустарники лишь с наиболее высокой частью низкого
приземного полога (0,4 – 0,5 м).
Во
всех упомянутых публикациях выделены группы птиц, предпочитающих различные
открытые пространства. По мнению одних авторов, это пойменные луга, в других работах
для тех же видов утверждается предпочтение степи, открытых болот или ерниковых
тундр. Независимо от ландшафтной и зональной принадлежности общим
для перечисленных территорий является относительная безлесность и в большинстве
случаев преобладание среднего по высоте травостоя, или кустарничков, низких
кустарников высотой 0,3 – 0,5 м или густого кочкарника такой же высоты
(расстояние между кочками менее 0,5 м), создающих хорошую укрытость для находящихся
под ними птиц малой и средней величины. Виды, приуроченные к подобным
условиям, выделены в топоархитектурную группу 11 – птиц, предпочитающих открытые пространства с низким (0,3 – 0,5 м
над землей) приземным пологом (причем образованным не обязательно только
растительностью). Соответственно все открытые пространства с низким приземным
пологом относятся к одноименному типу топоархитектуры.
Топоархитектураура
данного типа широко представлена в различных лугах, на
всевозможных травяных, низкокустарниковых или открытых кустарничковых болотах,
в кустарниковых или кустарничковых тундрах, а также среди сельскохозяйственных
полей, где культуры достигают указанной высоты.
Для
ряда видов, в отличие от предыдущих, прослеживается тяготение
к безлесным участкам с более низкой и редкой растительностью. Так, некоторые
авторы констатируют для них предпочтение лугов. Но это утверждение делается на
основе данных, полученных лишь в тех лугах, для которых характерна пастбищная
дигрессия растительности. По мнению других исследователей, работавших в
северных районах лесной зоны, те же виды предпочитают участки верховых болот с
очень редкими кустарничками. Общим для всех этих лугов и болот является преобладание
площадей с очень низкой растительностью (не выше 5 см) независимо от ее
видового состава. Кроме того, везде присутствуют низкие (не более 30 см) и
редкостойные кустарнички, или куртины злаков, или небольшие участки дернины,
или кочки, расстояние между которыми составляет 0,5 – 3,0 м (см. рис. 4). Птицы,
тяготеющие к подобным территориям, объединены в группу 12 – виды, предпочитающие
открытые территории без развитого
приземного полога с наличием редких куртин низкорослой растительности или
редких низких кочек. Соответственно все открытые пространства с
перечисленными характеристиками относятся к одноименному типу топоархитектуры.
Топоархитектура
данного типа представлена сравнительно широко. Она
может формироваться на опустыненных площадях, на солонцах, на лугах, чаще сухих,
или там, где высокий травостой по какой-либо причине деградировал. На низинных
травяных болотах она может присутствовать там, где проводится интенсивный
выпас, а также на осушенных участках, где болотная растительность деградировала,
а другая еще не развилась. В северных районах лесной зоны
такая топоархитектура широко представлена на верховых и переходных болотах в
основном там, где значительную площадь занимают мочажины, а по ним распределены
небольшие кочки или, к примеру, редкие кусты багульника. Кроме того, она
может формироваться и на безлесных буграх бугристых болот. В кустарниковых или
кустарничковых тундрах топоархитектура этого типа обычно представлена
там, где кустарники или кустарнички сильно изрежены и невысоки. Хозяйственная
деятельность в тундровой зоне приводит к формированию такой топоархитектуры
повсюду, где кустарники или кустарнички до этого создавали сравнительно сомкнутый
низкий полог. Чаще всего этому способствует интенсивный выпас оленей или
интенсивное движение гусеничной техники. Последний фактор может создавать
топоархитектуру данного типа даже на участках лишайниковых тундр, где совсем
могло не быть ни кочек, ни кустарничков. Роль последних элементов после интенсивных
транспортных воздействий начинают выполнять многочисленные куски дернины, вывороченные гусеницами машин.
Кроме
указанных в анализируемых публикациях выделены совокупности птиц, которые
тяготеют к территориям с наличием значительных участков с низкорослой растительностью
(не более 5 см) или совсем лишенных ее. Это, как правило, участки лугов, где травостой сильно поврежден,
пляжи, дно карьеров, отвалы песка, гидронамывы и т.п. Такие же условия
обитания часто формируются в лишайниковых тундрах и в местах песчаных раздувов.
По особенностям распределения эти птицы объединены в две группы видов,
предпочитающих соответственно топоархитектуру типов: 13 – открытые территории без развитого приземного полога; 14 – открытые территории, лишенные растительности.
Для птиц этих двух групп из-за особенностей их поведения развитая растительность
не создает укрытий, а, наоборот, является источником опасности (к примеру,
может служить укрытием охотящемуся хищнику).
Существует группа птиц, о предпочтении которых каждый из авторов сформулировал очень специфические определения (верховые болота; низинные болота; луга с кустарниками; склоны с выходами скал). Общим для всех упомянутых урочищ является то, что все они – открытые пространства с отдельно стоящими выделяющимися по высоте предметами. В качестве последних в одних случаях выступают деревья, высокие кусты, в других – скалы, столбы и т.д. Такие предметы служат присадами для наблюдения во время кормодобывания, токования или отдыха, а в некоторых случаях и местом устройства гнезда. Для разных видов заметна значимость различной величины данных предметов. Особенности гнездования этих видов позволили выделить три группы птиц и соответственно сформировать представления о трех типах топоархитектуры.
Так,
группа 15 объединяет виды, предпочитающие открытые
пространства с выделяющимися по высоте малыми предметами и элементами местности
(рис. 5, а).
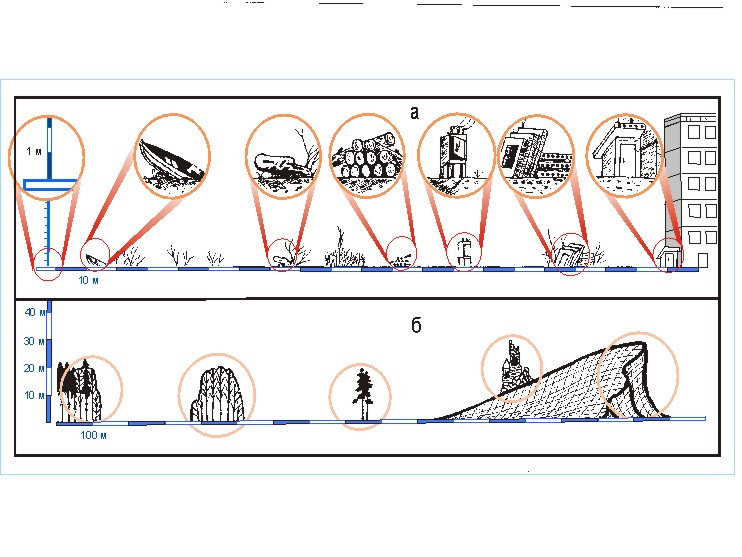
Рис. 5. Физиономические
профили топоархитектуры, сформированной открытыми пространствами.
а-
открытые пространства с отдельными выделяющимися по высоте малыми предметами; б – открытые пространства с отдельными
выделяющимися по высоте крупными предметами и элементами местности.
Эти предметы не более чем на 1 м возвышаются
над поверхностью земли. Примерами их могут служить крупные камни, кучи плавника,
различные предметы антропогенного происхождения. Расстояние между такими
предметами составляет 15 – 100 м. Обязательным условием, чтобы подобные пространства
соответствовали этому типу топоархитектуры, является отсутствие на большей
площади приземного полога. В группу 16 входят птицы, предпочитающие открытые пространства с выделяющимися по
высоте средними предметами и элементами местности (рис. 6). Для данного
типа топоархитектуры такие предметы должны на 1 – 5 м возвышаться над господствующей
растительностью. Это могут быть отдельные кусты, небольшие деревья, столбы,
заборы и т.п., расстояние между которыми составляет 15 – 50 м. В данном случае
для отнесения таких пространста к этому типу обязательно наличие на
преобладающей площади низкого приземного полога. Группа 17 объединяет птиц,
которые предпочитают открытые
пространства с выделяющимися по высоте крупными элементами местности (см.
рис. 5, б). Размеры таких элементов,
как правило, превышают 10 м по каждому измерению. В качестве примеров можно привести
отдельные группы высоких деревьев или небольшие перелески, лесные опушки,
скалы, крутые холмы, бугры с обрывами, большие здания, опоры высоковольтных
ЛЭП, вышки и т.д. В последнем типе топоархитектуры расстояние между крупными элементами
местности составляет, как правило, 100 – 1000 м.
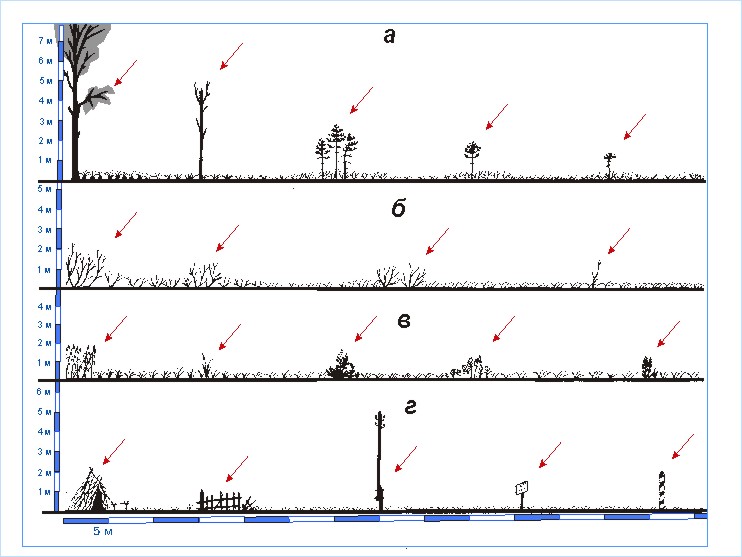
Рис. 6. Физиономический
профиль топоархитектуры типа “открытые пространства с отдельными выделяющимися
по высоте средними предметами и элементами местности”.
Выделяющиеся предметы: а
– деревья; б – кустарники; в – травы; г – антропогенные предметы.
Распределение
Riparia riparia, независимо от характера открытых
ландшафтов, сопряжено с близостью вертикальных обрывов в мягком грунте (в
гнездовой период наиболее активно используется открытое пространство не далее
1,5 – 2,0 км от обрывов). В связи с этим вид выделен в отдельную группу (18).
Выделяется также совокупность птиц, обитающих только на территориях с наличием крупных антропогенных сооружений, где постоянно
присутствуют люди (населенные пункты, промышленные установки, склады, автопарки
и т.п.). Эти виды объединены в группу 19, а пространства с соответствующими характеристиками
– в соответствующий тип.
Некоторые
птицы в одних классификациях характеризуются как предпочитающие водоемы, в
других – различные урочища суши. Последние урочища или как включения содержат
водоемы, или располагаются вблизи крупных водоемов или водотоков. С учетом этих
особенностей выделены группы птиц, предпочитающих чередование обширных открытых акваторий с малыми территориями: 20 –
занятыми рослым древостоем; 21 – свободными от древостоя и среднего
приземного полога (рис. 7). К этим типам топоархитектуры можно отнести
также прибрежные части любых крупных водоемов или водотоков вместе с узкой полосой
берега (100 – 200 м), если растительность там соответствует характеристикам
типа.
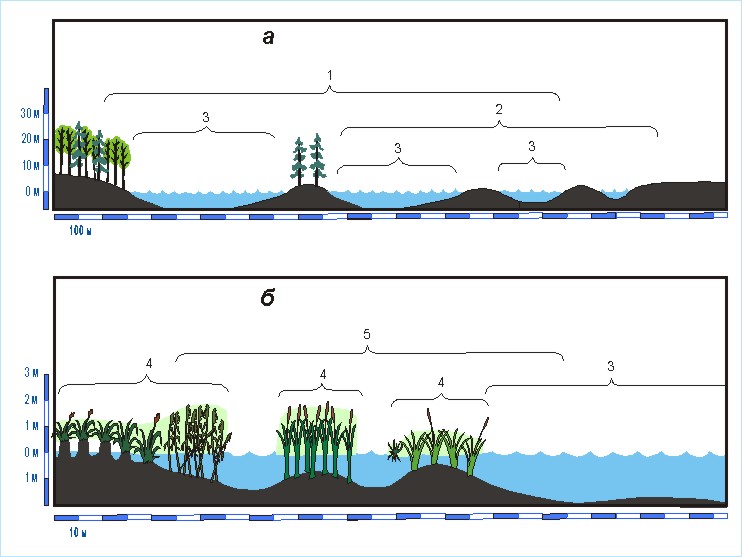
Рис. 7. Физиономические
профили топоархитектуры, сформированной с участием
акваторий.
1 – чередование обширных
акваторий и малых территорий, занятых рослым древостоем; 2 – чередование обширных акваторий и малых территорий, свободных от
древостоев и среднего приземного полога; 3
– открытые прибрежные пресноводные акватории; 4 – акватории, сплошь занятые надводной растительностью; 5 – чередование открытых акваторий и
занятых надводной растительностью.
Классификация
птиц водных пространств и их берегов [Юдкин, 1986б] позволила создать
представления о трех типах топоархитектуры: акватории,
сплошь занятые надводной растительностью; чередование открытых акваторий и занятых надводной растительностью,
создающей надводный полог средней высоты; прибрежные
пресноводные акватории без растительности (см. рис. 7). К последнему типу
следует относить акватории, расположенные не далее 1 км от ближайшей суши или
сплавины. Виды, обитающие на площадях с такими характеристиками, объединены в
соответствующие группы (22-24).
Кроме
того, каждый исследователь имеет свою специфическую трактовку относительно
предпочтения Tringa ochropus и Motacilla cinerea. Все же и
для этих птиц удалось выявить условия, общие в
совершенно в разных зонах и ландшафтах. Таким образом,
оба вида объединены в группу 25 – птиц, предпочитающих берега водоемов или водотоков среди полесья.
Здесь
приведены представления об идеальных типах топоархитектуры. В реальности один и
тот же участок может в разной степени соответствовать представлениям о нескольких
типах, а значит, на нем совместно обитают птицы, отнесенные к нескольким
топоархитектурным группам. В то же время некоторые площади по своим характеристикам
могут вообще не соответствовать никакому типу. Значительная
разномасштабность предлагаемых типов топоархитектуры не случайна. По размерам
тела отдельные виды птиц различаются в сотни раз, а
следовательно, и площади их индивидуальных участков также очень разные.
Известно,
что защитный фактор наиболее значим для птиц в период размножения, в остальное
время его роль снижается и из ведущих остается
трофический [Владышевский, 1974]. В результате в послегнездовое время происходит
значительное перераспределение птиц: они активнее начинают использовать
территории с меньшими защитными условиями [Юдкин, 1986а]. В
предложенной классификации группы сформированы по предпочтению территорий с определенной
топоархитектурой в период размножения.
1.3.3. Широтно-климатические группы птиц
В настоящее время пока не определен набор климатических параметров, непосредственно воздействующих на самих птиц или на факторы, лимитирующие их кормодобывание. Эта работа, возможно, будет выполнена в перспективе. Пока же приводится только классификация птиц по сходству их широтного распределения.
В
этом разделе из всех упомянутых публикаций анализируются только три [Вартапетов,
1984; Равкин, 1984; Юдкин, 1986б], поскольку в них представлен достаточный широтный
диапазон сравнимых между собой данных. Из всех параметров территориальных выделов,
характеризующих какой-либо класс, в данном случае заимствована только широтная
составляющая. В настоящее время имеется возможность сравнить полученные в этих
работах выводы со взглядами других исследователей,
сформировавшимися посредством иных методов [Брунов, 1978, 1980, 1982;
Кищинский, 1988]. В данной работе
пока не рассматриваются особенности распределения птиц в зависимости от
высотных изменений климата, так как классификация птиц выполнена для равнины.
Во
всех трех публикациях для северной части лесной зоны упоминаются две области
преференции: это предпочтение ряда урочищ северотаежной подзоны для одних видов
и нескольких урочищ из двух соседствующих подзон – северной и средней тайги –
для других. В указанных классификациях по видовому составу соответствующие
группы весьма сходны. Такой же характер обилия части этих
видов в Субарктике отмечен А.А. Кищинским (гипоаркты) и В.В. Бруновым (южнотундрово-лесотундровые
и северо-среднетаежные). Таким образом, по широтным особенностям распределения
отчетливо выделяется две совокупности видов: одна с высоким обилием на большей
части Субарктики, а другая на самом юге Субарктики и на самом севере умеренного
пояса.
Для
видов, распространенных в более южных районах лесной зоны, в анализируемых
классификациях выделены три полосы предпочтения: от средней тайги до подтаежных
лесов; южной тайги и подтаежных лесов и только южной тайги. Различия
этой части классификаций сводятся к тому, что материалы, полученные в долинах
Оби и Иртыша, показали предпочтение средней тайги в сочетании с более южными
подзонами для значительного числа видов (45% рассматриваемых в классификации).
Данные по Обь-Иртышским и Обь-Енисейским междуречьям позволили
выявить лишь 6% таких видов, а большинство остальных птиц списка
охарактеризовать как предпочитающие только южную тайгу. Различия результатов
трех классификаций можно легко объяснить характером выборок. Так, в междуречной
выборке, которую анализируют авторы, данные по высокопродуктивным
сомкнутым лесам имеются только из южной тайги. В результате для этой
выборки высокое обилие лесных птиц присуще только для южно-таежных проб.
Характерно,
что в другом варианте классификации, дополнительно использовав
данные по Приобью, Л.Г. Вартапетов отметил для тех же видов больший подзональный
спектр предпочтения. Результаты учетов в березово-сосновых лесах подтаежной
долины Тобола позволили выявить высокое обилие таежных видов и в этой подзоне
(в других работах в подтайге обследованы только мелколиственные леса). Как по
характеру широтного распределения, так и по видовому составу перечисленные
совокупности близки к выделенной В.В.Бруновым южно-таежной
географо-генетической группировке. Но анализ количественных данных показал, что
для указанных видов максимальное обилие характерно не только для южно-таежной подзоны. Небольшая часть птиц с одинаково
высоким (для данного вида) обилием распространена по всей северной половине
умеренного пояса, а другая, более многочисленная – примерно в средней его трети.
Как
для долин, так и для междуречий выявлена группа птиц, имеющих наибольшее обилие
в двух южных подзонах лесной зоны и, судя по публикациям [Равкин, 1973; Цыбулин,
1985; Торопов и др., 1985], они имеют столь же высокое обилие и гораздо южнее.
Примерно такими же широтными изменениями обилия В.В. Брунов характеризует выделенную
им южно-таежно-нетаежную группу.
Помимо
перечисленных, в двух вариантах опубликованных классификаций выделены группы
птиц, имеющих относительно высокое обилие в подтаежной
подзоне [Вартапетов, 1984; Юдкин, 1986б]. Так, если в Приобье подобных видов не
отмечено, то на более западном участке Обь-Иртышского
междуречья их обнаружено уже 17, а еще в более западной долине Тобола – 24.
Это, как правило, виды, основная часть ареала которых расположена южнее лесной
зоны. Увеличение их числа к западу в пределах подзоны, вероятно, обусловлено
тем, что северная граница ареалов многих из них при движении в том же
направлении смещается к северу аналогично границам климатических поясов.
Анализ
широтной компоненты этих классификаций показал, что в разных частях ареала
впечатление о связи видов с той или иной природной подзоной существенно меняется.
В то же время известны случаи массового заселения видом ранее не свойственной ему подзоны вместе с появлением там подходящих
местообитаний. Наиболее вероятно, что большинство видов вовсе не связано ни с
одной из выделенных современных природных зон или подзон. При наличии площадей
с пригодной топоархитектурой и субстратом лимитируют
широтное распространение видов исключительно климатические факторы.
По-видимому, это такие проявления климата, как продолжительность благоприятного
для размножения температурного периода, стабильность температур в это время,
отсутствие снежного покрова и т.п. Следовательно, для названий
широтно-климатических групп будут использованы особенности распределения птиц
относительно климатических поясов (но не принадлежность к какому-либо поясу). При этом для арктического и субарктического поясов названия
предлагаемых групп заимствованы из названий авифаун, выделенных А.А. Кищинским
[1988].
Таким образом, птиц, встреченных на территории лесной зоны
Западной Сибири, целесообразно разделить на следующие широтно-климатические
группы: 1 – эоарктический (наибольшее гнездовое обилие при прочих равных
условиях характерно для арктического пояса); 2 – гемиарктический (максимальное
гнездовое обилие возможно в пределах всего арктического и северной половины
субарктического поясов); 3 – гипоарктический (плотность их населения в период
размножения максимальна в субарктическом поясе); 4 – трансумеренный (виды на гнездовании широко распространены с
относительно высоким обилием в пределах большей части широт умеренного пояса);
5 – бореальный (обилие максимально в северной четверти умеренного пояса); 6 –
северный умеренный (обилие максимально в северной половине умеренного пояса) 7
– среднеумеренный (максимальное обилие представлено в средней трети умеренного
пояса); 8 – южный умеренный (обилие максимально в южной половине умеренного пояса).
Виды,
для гнездования которых благоприятные климатические условия (и, естественно, их
высокое обилие) находятся намного южнее анализируемой территории, за неимением
массового количественного материала пока не подразделены на группы и в классификации
условно объединены в девятую группу. При прочих равных условиях обилие
большинства видов к северу от выявленных зон предпочтения снижается
интенсивнее, чем при смещении к югу. Эта тенденция отмечена А.А. Кищинским
[1988] для северных птиц и подтверждается материалами по лесной зоне Западной
Сибири.
В
данной публикации не приводится типология климатической составляющей среды
обитания птиц. Это будет сделано в перспективе. Для формирования более
конкретных представлений о значимых совокупностях широтно-климатических
факторов и их типологии в настоящее время требуются дополнительные
трудоемкие исследования.
1.3.4. Долготные группы птиц
Классифицировать
виды птиц по долготным особенностям ареалов в настоящее время можно лишь
условно. Информация об этом аспекте распространения заимствована из сводок
"Птицы Советского Союза" [1951-1954], "Птицы СССР" [1982, 1987-1989]
и "Птицы России и сопредельных регионов" [1993].
При сопоставлении гнездовых ареалов с районами зимовок хорошо заметна сопряженность их долготных координат. То же прослеживается и для мест повторных отловов птиц, окольцованных на зимовках [McClure, 1974]. Известно, что многие птицы, районы гнездования которых лежат на иных долготах, чем районы их зимовок, при весенних перелетах сначала летят на север и лишь затем поворачивают к своим районам (к примеру, [Bruderer, 1997]). В связи с этим у таких видов отчетливо выражено запаздывание по долготе сроков прилета и гнездовых явлений [Фертикова, 2000].
Приведенные факты позволяют сформулировать гипотезу о том, что при прочих равных условиях, чем ближе долготные координаты гнездовой территории и мест зимовок, тем выше на этой территории гнездовое обилие, так как данные районы достигаются мигрантами быстрее и при меньших энергетических затратах. Это должно быть особенно актуально в тех случаях, когда численность вида лимитируется ресурсами на зимовках, а в пределах гнездового ареала ресурсы недоиспользуются. Для подтверждения изложенного следует напомнить, что снижение численности лесных птиц в умеренных широтах Северной Америки объясняется уничтожением лесов в районах их зимовок в Центральной и Южной Америке [Ricklefs, 1980; Robbins et al., 1989]. Для Европы также показано лимитирующее влияние зимовок на численность перелетных птиц в умеренных широтах [Бурский, 1993]. Поэтому при классификации видов по долготному распределению учтено не только положение гнездовых ареалов, но и районов зимовок.
В
результате по взаиморасположению районов гнездования и зимовок все виды (или
подвиды) птиц Палеарктики разделены на семь долготных групп: транспалеарктический,
крайнезападный, срединно-западный, западно-срединный, восточно-срединный, срединно-восточный,
крайневосточный.
В
транспалеарктическую группу объединены птицы, которые на гнездовании распределены
в большей части долгот Палеарктики, и зимовки их также обширны: простираются от
Африки до Юго-Восточной Азии. Крайнезападную группу составили птицы, зимующие в
Африке, а гнездящиеся в Западной и Восточной Европе. Максимальное их обилие на
гнездовании (при прочих равных условиях), вероятно, следует ожидать в секторе
20° з.д. – 30° в.д. Зимовки срединно-западных
видов занимают территорию от Африки или юга Западной Европы до Ирана или
Средней Азии. Гнездовой ареал большинства из них простирается от Западной Европы
до середины Красноярского края. Максимальное гнездовое обилие их следует
ожидать на долготах 0 – 60° в.д.
В
западно-срединную группу объединены виды, которые зимуют на пространстве от
восточной половины Африки до Индии. На гнездовании они распространены от
Центральной Европы примерно до долготы оз. Байкал. Максимальное обилие
предполагается в секторе 30 – 70° в.д. Восточно-срединные
виды зимуют на пространстве от Месопотамии до Бирмы. Гнездовой ареал
захватывает территорию от Восточной Европы до долготы р. Лены. Высокое
гнездовое обилие следует ожидать в пределах 70 – 100° в.д.
Срединно-восточные виды зимуют на территории от восточной части Индии до
Индокитая и Восточного Китая, а гнездятся от Западной Сибири до восточного побережья
Азии. Максимальное гнездовое обилие возможно в секторе 95 – 130° в.д.
Зимовки крайне-восточных птиц простираются от Индокитая до Австралии, островов
Индонезии, Филиппин, Новой Гвинеи и некоторых островов Океании, а гнездовые
районы, как правило, находятся на Дальнем Востоке и северо-востоке Сибири, а у
некоторых доходят до северо-западного побережья Северной Америки. Максимальное
обилие большинства из них вероятно в интервале 110 – 160° в.д.
Положение
186 видов птиц в каждой из четырех частей этой эколого-географической
классификации отражено в табл. 1. В примечании к
таблице перечислены названия групп каждой из четырех частей классификации, причем
они под теми же номерами, что в тексте и на рисунках.
1.4. Принципы оценки проявления факторов
Содержание
данного раздела не претендует на описание методики количественной оценки
проявления факторов. В его задачу входит лишь подробное разъяснение сущности
количественного проявления каждого из факторов и его характеристики. Для этого
в качестве примеров приводятся некоторые из возможных оценок. Технические же
стороны процедуры измерений в каждом конкретном случае должны разрабатываться
исследователем в соответствии с конкретными целями, необходимой точностью в
оценках и в зависимости от имеющихся возможностей.
Необходимость
в оценках проявления значимых для вида факторов может возникнуть как при
изучении его пространственного распределения, так и при прогнозировании его
обилия для необследованной территории. За основу характеристики распределения вида
i
внутри какого-либо обширного пространства наиболее рационально взять серию показателей
обилия на отдельных более мелких участках, на которые предварительно разделено
это пространство. Проявление значимых факторов часто бывает необходимо
определить отдельно для каждого такого участка n. Поскольку при изучении
или характеристике распределения вида i подобные участки n
принимаются за элементарные пространственные единицы, проявления на каждом из
них каждого значимого фактора обычно должно быть единой неделимой
характеристикой участка n.
1.4.1. Субстрат
Проявление
этого фактора может быть представлено как абсолютным его количеством, которое
можно выразить в единицах площади, так и относительным, выраженным
нормированным коэффициентом. Наиболее целесообразным показателем следует
считать относительное количество субстрата (его “концентрацию”) на определенной
площади. Это количество (ai) есть
отношение всей площади субстрата i (Si), представленного на оцениваемой площади территории
и/или акватории (Sx), ко всей
оцениваемой площади, т.е. ai=Si /Sx. Удобнее эти отношения выражать
в долях от единицы, что в дальнейшем будет показано при составлении модели.
Для
вида i
в репродуктивный период важно количество субстрата только на площади, занятой
топоархитектурой, которую он предпочитает. Следовательно, оценивать нужно
концентрацию субстрата по отношению лишь к той части площади участка n,
которая занята предпочитаемой топоархитектурой (Sx). Если исследователь работает сразу со многими видами
птиц, то при характеристике территории количество определённого субстрата
целесообразно оценивать не в целом для всей площади n, а дифференцированно
для каждой её части, занятой пригодной топоархитектурой. Зачастую и в пределах
площади с одной топоархитектурой могут отчётливо просматриваться участки, на
которых количество субстрата явно различается. В таком случае желательно дать
оценки для каждого участка отдельно.
Свободное воздушное пространство. Внутри этого типа
субстраты, пригодные для разных видов птиц, существенно различаются между собой
по многим характеристикам. Так, например, если для многих Accipiter spp. или Muscicapa
spp. пригодный субстрат – это не слишком большое пространство между кронами
соседних деревьев или высоких кустарников, то для кормодобывания многих Falco spp., Chlidonias spp., Hirundinidae
необходим гораздо больший свободный объём. Существенно различаются и
необходимые для разных видов высоты, на которых должно быть представлено свободное
воздушное пространство. Высота зависит от того, на каком уровне обычно проходит
кормодобывание определённого вида. Так, если для Saxicola spp. или Circus spp. свободное пространство необходимо
на высоте примерно от 0,5 м над землей или водой и выше, то для большинства
других видов данной субстратной группы свободное пространство ниже 5 м малозначимо,
и поэтому они могут найти для себя пригодные условия на большей площади. Такие
виды, как Apus spp., в основном
добывают корм выше яруса высоких деревьев. В результате для них благоприятный
субстрат представлен почти повсеместно.
Абсолютным
количеством субстрата этого типа следует считать площадь (Si), над которой на высоте
наиболее частого кормодобывания вида i и выше нет густых крон или
множества каких-либо иных высоких объёмных предметов. Относительным количеством
(или “концентрацией”) данного субстрата (ai) будет доля этой площади от всей площади оцениваемого
участка (Sx), т.е. ai=Si /Sx.
Незамкнутые вертикальные поверхности. Во многих
ситуациях к этому типу субстратов относятся лишь поверхности толстых стволов
деревьев и толстых ветвей. В таких случаях абсолютным количеством субстрата будет
сумма площадей поверхности всех участков стволов и ветвей, диаметр которых
превышает определенное значение. К примеру, для таких небольших по величине
птиц, как Dendrocopos minor или Sitta europaea, это критическое значение диаметра мало; для них вполне
пригодны поверхности стволов и ветвей толще 7-10 см. Для более крупных птиц
пригодными будут поверхности на гораздо более толстых частях стволов и ветвей.
Примерная
оценка абсолютной площади таких поверхностей (Si) не представляет особого труда: ее можно вычислить
через среднюю площадь пригодной поверхности на одном дереве или кусте (оценив
его высоту и диаметр на разных высотах) и среднее количество таких же деревьев
или кустов на единицу площади с пригодной топоархитектурой
(Sx).
Показатель относительного количества субстрата (ai) соответственно можно получить как ai =Si /Sx. Если на характеризуемой
площади к этому добавляются еще, к примеру, поверхности стен деревянных
строений или заборов, то несложно дополнительно оценить и их площадь. Сложнее
будет учесть все площади таких субстратов, как поверхности огромных камней,
скал, обрывов и т.п.
Замкнутые цилиндрические вертикальные и горизонтальные поверхности. Оценка
количества каких-либо субстратов из этих двух типов представляется наиболее
сложной. Так, абсолютное количество замкнутых поверхностей, ориентированных
вертикально, целесообразно выразить как сумму площадей проекций каждого их
элемента на вертикальную плоскость (рис. 8, а).
К примеру, для ситуации, изображенной на этом рисунке, сумма проекций всех элементов
такого субстрата (Si),
расположенного над площадью Sx, должна
быть рассчитана как Si=S1+S2+S3+S4+S5.
Следует уточнить, что при подсчете суммируются полные площади каждой из
проекций, учитывая и те их участки, которые совмещаются с другими проекциями.
Относительное количество (“концентрацию”) этого субстрата можно выразить
следующим образом: ai = (S1+S2+S3+S4+S5) /Sx.
Рис. 8. Схема,
иллюстрирующая оценку количества субстрата двух типов замкнутых цилиндрических
поверхностей.
а – вертикально ориентированные; б
– горизонтально ориентированные; S1 – S9 –
проекции каждого из имеющихся элементов субстрата; Sx – площадь,
для которой оценивается количество субстрата.
Сходным
образом оценивается и относительное количество субстратов, относящихся к типу
“замкнутые цилиндрические горизонтальные поверхности”. Но проецировать отдельные
элементы такого субстрата следует на горизонтальную плоскость (см. рис. 8, б). Характерно, что при таком
проецировании, чем ближе ориентация элемента субстрата к горизонтальной, тем
больше площадь его проекции. Для ситуации, изображенной на рисунке,
относительное количество субстрата будет выражаться как ai= (S6+S7+S8+S9) /Sx.
Некоторые
навыки для экспертной глазомерной оценки относительного количества субстратов
этого типа можно получить с использованием эталонных палеток. При их изготовлении
важны определенные соотношения ширины и густоты темных полос и белого поля.
Сетка, полученная таким образом, должна имитировать то изображение, которое увидит
эксперт на фоне неба, глядя снизу вверх на всю толщину слоя, в котором
представлен данный субстрат (вместе с наложением частей отдельных проекций друг
на друга). Для измерений необходимо иметь серию палеток, каждая из которых
имитирует изображение совокупности проекций при определенной концентрации
субстрата. В качестве примера на рис. 9 изображены палетки, соответствующие относительному
количеству субстрата 0,3; 0,5 и 0,7.
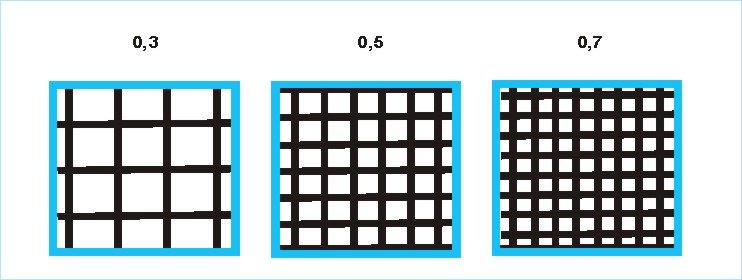
Рис. 9.
Эталонные изображения, имитирующие сумму проекций всех имеющихся элементов
субстрата типа “замкнутые цилиндрические горизонтальные поверхности”.
0,3, 0,5 и 0,7 –
относительное количество субстрата.
Сопоставляя
видимую снизу реальную картину сети пересекающихся элементов субстрата с
образцами сеток на палетках, можно визуально оценить, к какому из образцов
ближе реальная картина. При этом возникает определенная трудность. Так, если
сравнительно просто можно увидеть взаиморасположение элементов субстрата, находящихся
выше эксперта (кроны деревьев и высоких кустарников, переплетение проводов и
т.п.), то гораздо сложнее оценить густоту этих элементов, непосредственно у
поверхности земли (валежник, заломы жесткостебельного высокотравья, кроны
низких кустарников и т.п.). Необходимо уточнить, что при выполнении
количественной оценки такого субстрата следует учитывать лишь те его элементы,
которые пригодны для вида по диаметру поперечного сечения. При этом ясно, что
мелкие формы птиц могут использовать больший спектр элементов субстрата, чем
крупные. Для последних использование всей массы субстрата ограничено нижними
пределами поперечного сечения его элементов.
Все
выше перечисленное относится к экспертной оценке относительного количества
субстрата. Инструментальная оценка возможна с использованием крупномасштабных аэрофотоснимков.
В перспективе целесообразно разработать систему признаков для такого
дешифрирования снимков. Однако это не является целью данной книги.
Незамкнутые горизонтальные твердые и рыхлые поверхности. При оценке количества субстрата, относящегося к этим двум типам,
сложность заключается в дифференциации его на доступную и недоступную для птиц
части. Наиболее часто недоступным субстратом может быть поверхность
почвы. Возможность её эффективного использования птицами при кормодобывании
обычно ограничивается густой растительностью в приземном слое, в котором
находятся соответствующие птицы (рис. 10). В частности, слишком густой
травостой, низкие густые кустарники или кустарнички не позволяют птицам свободно
передвигаться при поиске и сборе корма. Особи многих видов могут регулярно использовать
толщу таких зарослей лишь в качестве убежища в момент преследования. В условиях
густого травостоя поверхность почвы или слежавшейся отмершей растительности
доступна для птиц в тех случаях, когда травостой создаёт густой слой выше деятельного
уровня определённого вида птиц, а под этим пологом имеется свободное пространство.
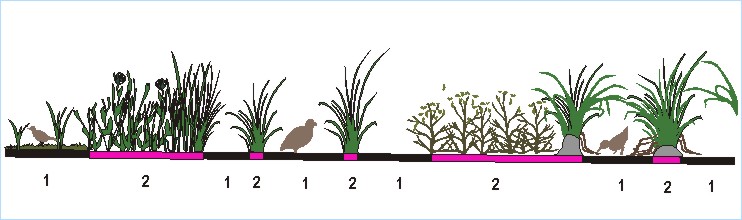
Рис. 10. Доступные (1) и недоступные (2) для кормового передвижения птиц части незамкнутой
горизонтальной поверхности.
Следует
обратить внимание на то, что одни и те же поверхности могут характеризоваться
различной доступностью для птиц разной величины. Так, в ситуации, когда травостой
смыкается верхними частями побегов, образуя густой слой в 0,7-1,0 м от земли, поверхность
почвы под ним доступна для птиц малой и средней величины. В то же время для
таких крупных видов, как Аnser spp. или Grus spp.,
находящаяся под этим пологом поверхность становится малодоступной. Другим
примером большей доступности поверхностей для мелких форм служит ситуация с
высоким и сравнительно густым кочкарником. В этих условиях средние и мелкие
птицы могут эффективно кормиться, передвигаясь по поверхности почвы (или
слежавшейся растительной ветоши) на участках между кочками. Для передвижения
крупных птиц близко расположенные друг к другу кочки создают серьёзные трудности.
В
качестве противоположного примера можно привести густой низкий травостой (20-30
см). Там, где он широко представлен, мелкие формы на значительной площади
обычно избегают добывать корм. При этом для свободного передвижения весьма
крупных птиц (многих Ciconiiformes,
Anseriformes, Galliformes, Gruiformes и т.п.) такой травостой не является
препятствием.
Кроме
того, разная доступность многих субстратов для птиц разной величины зависит ещё
и от характеристик самих поверхностей. Так, многие из субстратов, характеризующихся
для мелких птиц как сравнительно твёрдые поверхности, для средних и особенно
крупных видов представляют уже рыхлую или мягкую поверхность. Пример подобного
субстрата – плавающая на поверхности воды травянистая растительность. Она
обычно представлена или множеством широких плавающих на поверхности листьев
гидрофитов, или тонкой сплавиной из растительной ветоши. Такие мелкие птицы,
как, например, многие Motacillidae, кормящиеся в основном на твёрдых
поверхностях, легко передвигаются по этим субстратам. Из более крупных птиц,
часто кормящихся на плавающей травянистой растительности, почти все
приспособлены к мягким или рыхлым поверхностям (некоторые Rallidae, многие
Charadrii). В то же время более крупных птиц, из-за их значительной массы тела,
многие такие субстраты удержать не могут.
На
местности замеры примерного количества незамкнутых горизонтальных поверхностей
на обследуемом участке можно выполнять, по частным оценкам на серии малых
пробных площадок, в определённом порядке заложенных на характеризуемом участке.
Нужные измерения можно проделать также и на маршрутах, линии которых с разной
протяжённостью будут пересекать доступные и недоступные для изучаемого вида
площади. Но все перечисленные оценки будут экспертными. В подобных случаях
удобнее оценивать не абсолютную площадь доступных поверхностей (Si), а выполнять относительную
их оценку (ai=Si /Sx). Во многих случаях для
пространств, свободных от сомкнутых деревьев и высоких кустарников, возможен
инструментальный способ оценки. Для этого вполне пригодна информация,
представленная на крупномасштабных аэрофотоснимках (М 1:1000 – 1:5000). Признаками
относительного количества доступных поверхностей должна послужить тональность и
текстура различных участков изображения. Их можно выделять с помощью ЭВМ, что
минимизирует субъективизм исследователя. Такие оценки могут выполняться для
значительных площадей.
Сходным
образом характеризуется доступность и количество субстратов, относящихся к типу
“поверхность
воды”. Абсолютное количество субстратов типа “толща воды” (Si) оптимально оценивать как
площадь акватории, под которой располагается определенная глубина. При этом
доступной частью такого субстрата следует считать площадь акватории (Si), свободную
от густой не только надводной, но и подводной растительности. Относительное
количество этого субстрата определяется по отношению ко всей площади характеризуемого
участка (Sx), независимо
от того, представлена она только акваторией или ещё и территорией (ai=Si /Sx). Субстраты двух последних
типов весьма легко распознаются не только на аэрофотоснимках, но и на
космических снимках высокого разрешения.
1.4.2. Топоархитектура
Абсолютным
количеством топоархитектуры на участке n является площадь, на которой она
представлена. Но более удобен относительный показатель количества: доля этой
площади от общей площади участка n. В дальнейшем относительный
показатель, как и количество субстрата, будет выражаться в долях от единицы.
При
количественной оценке представленности какой-либо топоархитектуры на участке n
основная задача заключается в определении – какие
площади по своим условиям соответствуют характеристикам данной топоархитектуры.
При таких оценках непосредственно на местности это соответствие может быть
установлено лишь экспертно. Инструментально оценить количество топоархитектуры
можно с помощью аэрофотосъёмки. Топоархитертура большинства типов сравнительно
легко распознается по прямым признакам на снимках масштаба 1:10000 и крупнее.
Весьма удовлетворительные результаты можно получить и при чтении снимков М
1:20000 – 1:25000, но лишь по отдельным типам топоархитектуры (различные
древостои, полесье, лесополье, средний полог среди открытых пространств). При
работе со снимками площади, занятые определённой топоархитектурой, можно выделить
по серии специально разработанных формализованных признаков (определённые тон,
плотность, текстура фотоизображения и т.п.), оконтуривать их и выполнять замеры
оконтуренной площади. Некоторую информацию можно получить при сопоставлении
космических снимков высокого разрешения (к примеру, МСУ-Э), сделанных в разное
время и по разным каналам.
При
характеристике участка n на пригодность для обитания на нём
всех возможных видов птиц дифференцированно выделяются и оконтуриваются площади
для каждой присутствующей там топоархитекты. При этом не следует забывать, что
одна и та же площадь по своим свойствам может соответствовать характеристикам
сразу нескольких разновидностей топоархитектуры. Так, участок сомкнутого
рослого древостоя у лесной опушки соответствует топоархитектуре «сомкнутый
рослый древостой», а так же часть этого древостоя на глубину нескольких десятков
метров от опушки будет ещё составной частью топоархитектуры “полесье” (см. рис.
3, г). На меньшую глубину
приопушечная полоса этого древостоя будет ещё частью “лесополья”. Кроме того,
лесная опушка вместе со значительной частью соседнего открытого пространства по
своим характеристикам соответствует также топоархитектуре “открытые
пространства с выделяющимися по высоте крупными предметами и элементами
местности” (см. рис. 5). При переходе от рослого древостоя к открытому
пространству в узкой приопушечной полосе часто встречаются отдельные кусты.
Они, совместно с некоторыми низкими ветвями крайних деревьев, могут выполнять роль выделяющихся по высоте средних предметов (см.
рис. 6, а). Другой пример наложения разных топоархитектур (особенно разного масштаба) друг на
друга приведён на рис. 7.
1.4.3. Широтно-климатические факторы
Проявление
широтно-климатических факторов на участке n удобнее всего выразить в величине
отклонения климатических условий от тех, в которых вид i приспособлен
эффективнее всего размножаться. Это отклонение должно рассчитываться как
разница значимых климатических характеристик в зоне предпочтения и на участке n.
Например, наиболее благоприятным для гнездования какого-либо вида является
период с определёнными температурными и другими метеоусловиями продолжительностью
ti дней. На
участке n продолжительность периода с такими условиями составляет tn дней. Таким образом, проявление
широтно-климатического фактора на участке n равно cni = ti -tn и будет выражено в количестве
дней.
Но,
как уже упоминалось, пока ещё отчетливо не выявлены те климатические параметры,
которые непосредственно определяют широтное и высотное распространение видов.
Для Западно-Сибирской равнины установлены лишь координаты зон
широтно-климатического предпочтения многих видов. Судить об отклонениях климатических
условий на участке n от тех условий, которые благоприятны для вида, в данный
момент приходится по расстоянию участка n от зоны предпочтения. Причем
делать это приходится с определенными оговорками, хотя на этой равнине более, чем где-либо, климатические параметры сопряжены с
широтными координатами. Удалённость участка n от зоны предпочтения
можно выразить как в градусах широты, так и в километрах. Однако за пределами
Западно-Сибирской равнины изменения климатических условий уже нельзя считать функцией
расстояния по широте.
1.4.4. Удалённость от долготного сектора предпочтения
По
определению, это – фактор расстояния. Он показывает, насколько участок n
удалён от долготы района зимовок вида i. Данный фактор наиболее удобно
выражать в километрах. Не следует удалённость выражать в градусах долготы, так
как на разных широтах один такой градус составляет разное линейное расстояние.
Глава 2. ФОРМИРОВАНИЕ ОБИЛИЯ ВИДА НА ОПРЕДЕЛЕННОЙ ПЛОЩАДИ
2.1. Ключевые понятия
В гл. 1 изложены представления о системе
факторов среды, определяющих пространственное распределение птиц. Для того
чтобы понять, как эти факторы влияют на распределение птиц, необходимо уточнить
и расширить традиционный понятийный аппарат зоологов и зоогеографов.
Большинство понятий, которые используются в настоящей работе, – общепринятые, а
термины, обозначающие их, как правило, имеют однозначное толкование.
Сравнительно новыми, традиционно не используемыми понятиями, являются: благоприятный
для вида субстрат, предпочитаемая топоархитектура, элементарный пространственный
выдел, идеальная и реальная плотности вида, характер пространственной
неоднородности обилия вида. Первые две из перечисленных категорий разъяснялись
в предыдущей главе. В данном случае остановимся на оставшихся четырех.
Элементарный пространственный
выдел. В географии при характеристике континуальных
объектов или явлений часто встает проблема наименьшей единицы рассмотрения
[Харвей, 1974]. Применительно к характеристике распределения птиц это выглядит
следующим образом. Население любого вида i на какой-либо обширной площади неоднородно по
плотности, а плотность представляет собой пространственный континуум. Изучение
закономерностей пространственного распределения вида по этой территории, а
зачастую и характеристику такого распределения рационально выполнять на основе
сравнения показателей обилия вида на более мелких частях данной площади. Для
этого необходимо предварительно разбить всю характеризуемую площадь на мелкие
части. Причем в зависимости от поставленных задач может быть несколько рангов
такого деления. Самым дробным должно стать разделение характеризуемой обширной
площади на элементарные (для конкретного этапа рассмотрения) участки.
Показатель обилия вида рассчитывается в целом для каждого участка, а
неоднородность обилия вида внутри участка не рассматривается. Каждый участок
будет именоваться элементарным пространственным выделом и будет наименьшей
единицей рассмотрения пространства. Совокупность особей вида i в каждом выделе будет
наименьшей единицей рассмотрения населения этого вида. Таким образом, под
элементарным пространственным выделом в данном случае следует понимать целостный
в пространстве и в дальнейшем неделимый участок территории или акватории, на котором
определяется количество особей определенного вида или комплекса видов для анализа
или характеристики пространственных изменений этих величин.
Элементарные пространственные выделы в
идеальном случае должны быть равны по форме и величине, которые определяет
предметник сообразно конкретным задачам и исходя из свойств изучаемого вида. На
практике в подавляющем большинстве случаев приходится ограничиваться
характеристикой обилия не на всех таких единицах, а лишь на очень ограниченной
выборке. Более того, такие выделы могут различаться между собой по форме и
величине.
Изучение или характеристика
пространственного распределения вида начинается со сравнения и анализа показателей
его обилия в элементарных выделах. Затем на основе выявленных закономерностей
можно от дробного рассмотрения переходить к описанию на разных уровнях
обобщения. При таком переходе можно объединять элементарные выделы в выделы
большего ранга, оконтуривая их на карте на основе
полученных знаний о закономерностях распределения вида.
Под характером территориальной неоднородности обилия понимается вся совокупность пропорций (матрица), в
которых соотносятся между собой показатели обилия в пределах определенной
выборки (набора обследованных территориальных выделов).
Идеальная плотность вида или подвида (Ni0). Для того
чтобы судить о плотностном потенциале вида i в гнездовой период и соответственно о возможных значениях
его обилия при тех или иных проявлениях факторов, необходимо ввести понятие
идеальной плотности. В предлагаемой концепции данная характеристика должна
стать одним из ключевых понятий. Итак, Ni0 – это
плотность населения вида (или подвида), которая формируется в гнездовой период
на площади, сплошь занятой пригодной для него топоархитектурой при
одновременном выполнении пяти условий: в наиболее благоприятных для вида
широтно-климатических условиях; на минимальном расстоянии от мест зимовок (для
перелетных видов на равнине при соблюдении предыдущего условия это расстояние в
идеальном случае минимально на долготе места зимовок); при наиболее
характерном для большинства естественных экосистем интервале концентрации
кормов; при определенном эталонном количестве субстрата; при отсутствии
лимитирующего воздействия других факторов. За эталонное количество субстрата
принята величина 0,6, выбор которой будет объяснен в следующей главе.
Подробнее
сущность этой категории заключается в следующем. В период размножения птица
(или гнездовая пара) привязана к ограниченной территории, на которой находится
ее гнездо или менее мобильный, чем она, выводок. Следовательно, она вынуждена
обитать на относительно постоянной ограниченной площади (не следует путать с
понятием гнездового участка). Минимальная достаточная для нее величина этой
площади (si) определяется
соотношением, с одной стороны, потребности данной особи (или пары) в каком-то
количестве корма, необходимом для нормальной ее жизнедеятельности и роста ее
потомства (vi, ккал), с
другой стороны, концентрации (или интен сивности возобновления) доступных кормовых
ресурсов (vn, ккал/км2). Таким образом, si=ks
vi
/vn.
Количество таких минимальных достаточных площадей, которое может поместиться на
единице площади, сплошь занятой предпочитаемой топоархитектурой, в идеале и
должна характеризовать величина Ni0, т.е.
Ni0 = kn sb
/si , (1)
где ks и kn – коэффициенты; sb – площадь, занятая пригодной
топоархитектурой.
Из
общепринятых понятий плотности к идеальной наиболее близка оптимальная
плотность популяции. Но идеальная плотность вида – это оптимальная плотность
популяции в упомянутых идеальных условиях.
Основное
свойство Ni0 – ее зависимость от средней величины одной особи
вида. Такая зависимость обусловлена следующим. Известно, что количество требующегося
кормового ресурса определяется метаболизмом птицы, который, в свою очередь,
находится в прямой зависимости от массы ее тела [Lasiewski, Dawson, 1967; Гаврилов,
1977; Kendeigh et al., 1977; Шмидт-Ниельсен, 1987]. Можно предположить, что в
сходных условиях для птиц с одинаковой интенсивностью метаболизма и
потребляющих одни и те же корма величина достаточных участков находится в
прямой зависимости от массы их тела. Следовательно Ni0
, согласно уравнению (1), находится в обратной зависимости от этой массы.
Внутри одного вида величины достаточных минимальных участков отдельных особей
должны различаться столь же незначительно, как и размеры
их тела. В результате Ni0 – величина постоянная
для вида в пределах всей территории с перечисленными идеальными условиями.
Такие предположения могут вызвать возражения у исследователей, работающих с эмпирическими данными по обилию птиц. Чтобы это предупредить, необходимо напомнить, что изложенное относится не к традиционным показателям обилия (в пересчете на всю площадь любого обследованного выдела), а к идеальной плотности населения вида. Даже для традиционного обилия можно обнаружить связь с величиной одной особи вида, хотя и не столь однозначную [Blackburn et al., 1990]. Однозначно обратная зависимость обилия вида от величины одной его особи отчетливо прослежена, например, у беспозвоночных [Гиляров, 1944]. На основе данных по птицам, хотя и косвенно, также можно констатировать ее наличие. Так, рассчитанные по эмпирическим материалам уравнения регрессии иллюстрируют прямую сопряженность величины гнездового (или кормового) участка вида с массой тела одной его особи [Shoener, 1968; Harrestad, Bunnell, 1979; Baker, Mewaldt, 1979, цит. по Дольник, 1993; Гаврилов, 1988]. В то же время величины гнездового и кормового участков являются одним из механизмов, ограничивающих верхний уровень обилия вида.
Реальная (имманентная) плотность вида или подвида (Nint). При оценках численности популяции, особенно когда стоят задачи по изучению ее многолетних изменений, или при анализе ее географической неоднородности простые сравнения обилия в выделах или их совокупностях зачастую бывают некорректными. Как правило, не вся площадь выдела n, в котором вид i встречен, бывает пригодна для его обитания. Обычно какие-либо ее части вообще не посещаются этими животными. Следовательно, показатель обилия вида в конкретном выделе помимо численности популяции в данный год или в данном районе отражает еще и количество пригодной для вида площади в этом выделе. Исключить влияние указанного фактора можно, сравнивая обилие вида только в одинаковых по условиям обитания выделах, что на практике сделать весьма трудно, или пересчитав обилие не на всю площадь n, а лишь на пригодную ее часть. Последний показатель и будет в дальнейшем именоваться реальной или имманентной плотностью вида i в выделе n (Nint).
Таким образом, реальная (имманентная) плотность вида i в выделе n – это количество птиц встреченных в выделе n в пересчете на единицу площади лишь той его части, которая занята пригодной для данного вида топоархитектурой, т.е.
Nint =Nin emp /bin , (2)
где Nin emp – эмпирический показатель обилия вида i в выделе n, bin – доля площади с пригодной топоархитектурой.
В отличие от предыдущей категории, реальная плотность – всегда реальное число особей, и оцениваться эта величина может для любого времени и при любых проявлениях остальных значимых факторов, а не только в гнездовой период и при прочих максимально благоприятных условиях.
2.2. Аксиоматика
Концептуальная
модель, которая должна иллюстрировать принципиальный характер зависимости
обилия птиц в репродуктивный период от совокупного влияния значимых факторов
среды, будет основана на следующих аксиоматических утверждениях и следствиях из
них. Каждое из этих утверждений делается для ситуации, когда все прочие условия
равны.
1.
Вид i в природе
может обитать и успешно размножаться лишь на площади, которая занята
предпочитаемой им топоархитектурой.
2.
Количество особей вида i в выделе n пропорционально возможному
количеству пригодных для вида участков, достаточных по площади для обитания особи
(пары или выводка).
3.
Возможное количество достаточных для вида i участков обитания в выделе n пропорционально той ее площади, на которой представлена
предпочитаемая этим видом топоархитектура.
4.
Возможное количество достаточных участков в выделе n обратно пропорционально их величине.
5.
Величина достаточных участков обратно пропорциональна ресурсу доступных кормов.
Доступность кормов, в свою очередь, положительно сопряжена с количеством
соответствующего субстрата. Поэтому, чем больше количество соответствующего субстрата
на территории с пригодной топоархитектурой, тем меньшая площадь такого участка
требуется для обитания особи (пары или выводка).
6.
Чем ближе на территории выдела n
климатические условия к тем, которые предпочитает вид i, тем большая доля таких возможных
участков обитания будет занята.
7.
Чем ближе выдел n к месту,
где вид i может
пережить самый неблагоприятный период года (в данном случае – место зимовок),
тем больше доля таких пригодных участков обитания в этом выделе будет занята.
2.3. Концептуальная модель зависимости обилия от внешних факторов
Гиперобъем
реализованной ниши вида i
ограничен участками осей параметров ниши.
Как уже указывалось, группы параметров ниши подобраны таким образом, чтобы их
оси были ортогональными. Поэтому и стороны условной фигуры, ограничивающей
гиперобъем реализованной экологической ниши вида i, также будут ортогональными. Это
позволяет рассчитать гиперобъем фигуры как простое произведение ее сторон.
Основываясь на этом и на изложенных в предыдущем разделе утверждениях,
концептуальную модель зависимости обилия птиц от совокупного воздействия
значимых факторов в самых общих чертах можно представить следующим образом:
Nin=Ni0 f(ain)
f(bin) f(cni) f(dni) k1..z
или
(3)
lnNin=lnNi0+lnf(ain)+lnf(bin)+lnf(cni)+lnf(dni)+lnk1,...,z ,
где Nin – обилие вида (подвида) i в гнездовой период в выделе n; Ni0 – идеальная плотность вида n; f – функция обилия от факторов среды; ain – количество пригодного для вида (подвида) i доступного субстрата на площади, занятой предпочитаемой топоархитектурой в выделе n; bin – доля площади выдела n с топоархитектурой, которую предпочитает вид (подвид) i; cni – отклонение широтно-климатических условий в выделе n от их интервала, предпочитаемого видом i (или удаленность выдела n от широтно-климатической зоны предпочтения вида); dni – удаленность выдела n от долготного сектора предпочтение вида i; k1,...,z – функция обилия от внутренних и неучтенных внешних факторов (они должны быть выделены из всей совокупности условий таким образом, чтобы каждый из них не был функцией какого-либо другого).
Основное
назначение этого уравнения – выполнять познавательные функции. Во-первых, оно
помогает понять суть явления распределения животных в пространстве, во-вторых,
способствует дальнейшему познанию данного явления. Кроме того, оно может быть
основой для формирования практической модели прогнозирования обилия на
какой-либо территории. Такой прогноз может быть использован не только для
оценки полноты знаний, но и для прикладных целей.
Таким
образом, для удобства дальнейшего использования этой модели целесообразно,
чтобы все имеющиеся там функции от факторов были представлены нормированными
коэффициентами, выраженными в долях от единицы. Тогда произведение этих функций
(без умножения на идеальную или реальную плотность вида) будет иллюстрировать
лишь характер пространственной неоднородности обилия. Размерностью обилия
(число особей на единицу площади) в данной формуле обладает лишь Ni0. Однако
реальная плотность в условиях, определенных для идеальной плотности, в силу
ряда причин зачастую бывает ниже идеальной (к примеру, из-за чрезвычайно
высокой гибели на зимовках в отдельные годы, из-за существенного сокращения
пригодных площадей на зимовках при одновременном изобилии их в местах гнездования
и т. д.). Следовательно, для каких-либо практических расчетов в уравнение (3)
на место Ni0 необходимо
будет подставлять реальную плотность, определенную в ограниченной выборке
эмпирическим путем и пересчитанную к идеальным условиям.
Глава 3. ХАРАКТЕР ЗАВИСИМОСТИ ОБИЛИЯ ВИДА ОТ ЗНАЧИМЫХ ФАКТОРОВ
В предыдущей главе предложена концептуальная модель совокупного влияния факторов среды на обилие птиц (см. формулу 3). Однако это уравнение лишь в самых общих чертах иллюстрирует основные зависимости. В то же время имеющиеся количественные данные по обилию птиц в значительном количестве территориальных выделов и информация об условиях их обитания уже сейчас позволяют существенно детализировать эту модель. В настоящей главе на основе анализа имеющихся эмпирических материалов решаются три задачи:
1) вывести уравнение регрессии идеальной плотности вида от средней массы одной особи;
2) получить идеализированные уравнения зависимости обилия вида от значимых факторов среды;
3) составить развернутую модель зависимости обилия птиц от совокупного воздействия этих факторов.
Необходимо отметить, что оценить коэффициенты регрессии сразу по всем факторам среды позволяет множественная регрессия. Но такая оценка не позволит проиллюстрировать сущность явления организации распределения птиц, а значит, затруднительно будет сформировать идеализированные представления об этом.
3.1. Используемый материал и способы его анализа
Для
решения поставленных задач использованы результаты учетов птиц в различных
районах Западной Сибири. Эти материалы собранны автором лично и совместно с
Л.Г. Вартапетовым, В.Г. Козиным, К.В. Тороповым, Е.Л. Шором, Б.Н. Фоминым и
В.М. Ануфриевым, которым автор выражает глубокую признательность. Сведения по обилию птиц получены в результате маршрутных учетов с
последующим пересчетом данных на площадь по среднегрупповым дальностям обнаружения
[Равкин, 1967] в период с 16 мая (или с 1 июня) по 15 июля 1981-1992 гг. Всего
использованы характеристики обилия 186 видов птиц в 164 территориальных выделах
(в ранге ландшафтных урочищ) с 11 ключевых участков, распределенных в интервале
57 – 69° с.ш. и 65 – 84° в.д. В
различной степени представлен материал из каждой природной подзоны от
подтаежных лесов до субарктических тундр Западной Сибири. Данные по
встречаемости и характеру пребывания птиц, полученные в этих исследованиях,
опубликованы [Бобков и др., 1997; Юдкин и др., 1997; Вартапетов, Юдкин, 1999].
Проявления
факторов количества субстрата и представленности топоархитектуры в каждом
территориальном выделе (в данном случае на трансекте) получили экспертную
балльную оценку. Их количественные оценки на местности выполнялись лишь на трех
ключевых участках, причем на одном из них не во время исследования птиц, а по
прошествии пяти лет. Для остальных районов работ оценки выполнены экспертно в
камеральных условиях с привлечением фотоснимков, сделанных на трансектах. Кроме
того, частично необходимая информация считывалась с космических снимков
высокого разрешения (МСУ-Э). Поскольку оценки факторов во многих выделах выполнены
по прошествии иногда значительного времени после учета птиц, в некоторых
случаях не исключены погрешности. Этот недостаток частично компенсировался
большим объемом использованного эмпирического материала. Проявление каждого из
этих двух факторов получило нормированную оценку в долях от единицы.
Любые
современные методы оценки обилия значительной части видов
птиц грубы. Поэтому выявление характера изменений обилия по градиенту
каждого из факторов и значений Ni0
ограничивалось графическим способом. Стандартная функция, наиболее удачно
аппроксимирующая эти изменения, подбиралась экспертно с таким расчетом, чтобы
отражать не особенности конкретного вида в определенной подвыборке, а то общее,
что повсеместно характерно для изменений обилия птиц по данному фактору.
Коэффициенты регрессии уравнений рассчитывались способом наименьших квадратов.
При этом для Ni0 и f(ain) задача значительно упрощалась логарифмированием данных.
Для
каждого из расчетов использовался не весь материал, а лишь данные по тем видам
и тем пробам (обследованным выделам), которые в каждом конкретном расчете отвечали
следующим требованиям. Так, для расчета уравнения регрессии Ni0 от средней
массы одной особи вида, согласно определению идеальной плотности, брались не
эмпирические показатели обилия, а значения реальной (имманентной) плотности
вида (Nint). Как уже указывалось в гл. 2, этот показатель
получен следующим образом:
Nint=Nin emp / bin,
где Nin emp – эмпирический показатель обилия вида i в гнездовой период в выделе n
(не следует путать с плотностью гнездования); bin – площадь
выдела n, которая
занята предпочитаемой видом i топоархитектурой.
Кроме
того, согласно определению Ni0 , для анализа ее
зависимости от массы тела одной птицы из оставшихся подвыборок отобраны пробы с
количеством субстрата 0,5 – 0,7 и только те, которые относятся к
широтно-климатической зоне и долготному сектору предпочтения анализируемого
вида. Из рассмотрения исключены колониальные птицы из-за
высокой агрегированности их распределения, дуплогнездники, поскольку их гнездовое
обилие гораздо сильнее, чем других видов, лимитируется количеством
гнездопригодных мест: дупел или искусственных гнездовий. Кроме того,
исключены виды, для которых пригодными субстратами являются поверхность воды и
воздушный объем (количественная оценка этих субстратов в данной работе наименее
корректна).
Не только для данного расчета, но и повсеместно, где
использовалась характеристика Nint, из выборки исключены
все пробы, в которых доля площади с предпочитаемой видом топоархитектурой
составляет 0,1n и менее. Это сделано по следующим
соображениям. Количество особей вида i в выделе n мало, если там мала
площадь, занятая соответствующей ему топоархитектурой.
Как следствие этого малό и количество встреч птиц вида i
во время его учета. Вычисленные по малому количеству встреч показатели обилия
весьма ненадежны [Равкин, Челинцев, 1990]. Надежность таких значений
уменьшается еще на порядок и более при расчете Nint (за счет
деления на такую малую величину, как 0,1 и менее).
В
результате всех перечисленных ограничений для формирования представлений о
величинах Ni0 остались данные по 37 видам. Для каждого из них
по отобранным пробам рассчитана одна средняя величина Nint за все
годы наблюдения. Таким образом, полученные значения по возможности максимально
приближены к средним многолетним. Данные о средней
массе одной взрослой особи заимствованы из сводки “Птицы Советсткого Союза”
(1951-1954). Результаты расчетов приведены в табл. 2.
Т а б л и ц а 2
Характеристики реальной (имманентной) плотности видов в идеальных условиях
|
Вид |
Средняя масса одной
особи (m), г |
Средняя реальная плотность (Nint), особей/км2 |
Суммарная
биомасса Nint количества
птиц, кг/км2 |
Энергия,
трансформируемая Nint количеством
птиц, тыс. ккал/(сут. км2) |
n |
|
Lyrurus tetrix |
970 |
7 |
7,5 |
1,4 |
3 |
|
Tetrao
urogallus |
3085 |
4 |
10,6 |
1,4 |
3 |
|
Tetrastes
bonasia |
382 |
28 |
12,4 |
3,3 |
1 |
|
Grus grus |
5475 |
1 |
5,2 |
0,5 |
2 |
|
Porzana porzana |
86 |
24 |
2,4 |
1,2 |
2 |
|
Pluvialis
apricaria |
205 |
21 |
4,2 |
1,5 |
3 |
|
Tringa glareola |
60 |
78 |
4,7 |
2,8 |
4 |
|
T,nebularia |
205 |
20 |
3,3 |
1,3 |
2 |
|
T.stagnatilis |
59 |
50 |
3,5 |
2,0 |
1 |
|
Philomachus
pugnax |
131 |
38 |
5,2 |
2,2 |
3 |
|
Gallinago
gallinago |
100 |
37 |
4,0 |
1,9 |
2 |
|
Limosa
lapponica |
263 |
22 |
5,9 |
1,9 |
2 |
|
Colunba oenas |
280 |
6 |
1,7 |
0,5 |
2 |
|
Streptopelia
orientalis |
180 |
11 |
2,4 |
0,9 |
3 |
|
Cuculus canorus |
107 |
19 |
2,1 |
1,0 |
2 |
|
Alauda arvensis |
38 |
80 |
2,9 |
2,4 |
2 |
|
Anthus
trivialis |
23 |
117 |
2,6 |
2,6 |
2 |
|
An. cervinus |
20 |
117 |
2,3 |
2,4 |
2 |
|
Motacilla flava |
17 |
178 |
2,8 |
3,3 |
6 |
|
Oriolus oriolus |
73 |
37 |
2,8 |
1,7 |
1 |
|
Perisoreus
infaustus |
93 |
11 |
0,9 |
0,5 |
3 |
|
Bombycilla
garrulus |
56 |
22 |
1,2 |
0,8 |
2 |
|
Locustella
lanceolata |
13 |
160 |
1,9 |
2,5 |
2 |
|
Acrocephalus
schoenobaenus |
12 |
240 |
2,9 |
3,7 |
2 |
|
Ac. dumetorum |
14 |
175 |
2,1 |
2,7 |
3 |
|
Sylvia borin |
17 |
150 |
2,7 |
2,9 |
1 |
|
S. communis |
16 |
183 |
2,9 |
3,4 |
2 |
|
Phylloscopus
trochilus |
9 |
178 |
1,6 |
2,3 |
4 |
|
Ph. borealis |
11 |
150 |
1,5 |
2,1 |
1 |
|
Luscinia
svecica |
17 |
151 |
2,6 |
2,9 |
2 |
|
Turdus
atrogularis |
105 |
11 |
1,0 |
0,6 |
1 |
|
T. philomelos |
74 |
17 |
1,2 |
0,8 |
2 |
|
Parus major |
18 |
124 |
2,5 |
2,6 |
1 |
|
Fringilla
coelebs |
22 |
127 |
2,8 |
2,8 |
2 |
|
Carpodacus erythrinus |
22 |
106 |
2,3 |
2,3 |
2 |
|
Emberiza pusilla |
14 |
180 |
2,5 |
3,1 |
4 |
|
Em. aureola |
23 |
120 |
2,6 |
2,6 |
1 |
Линейный
характер зависимости обилия Nin emp
от bin
(что будет показано в следующем разделе) делает корректным использование Nint
в подобных сравнениях. Следует отметить, что для каждого вида с надежными оценками
обилия значения Nint , рассчитанные для разных урочищ, оказались
весьма близкими между собой. Естественно, это сходство проявляется при
различных bin , но при прочих равных условиях. К примеру, в
пределах одного ключевого участка для всех территорий с ain=0,5 – 0,7
коэффициенты вариации значений Nint не превышали 35%.
Характер
изменений обилия по внешним факторам анализировался отдельно для каждого вида.
Для объективного выявления этого характера по каждому фактору следует
рассматривать только те изменения обилия, которые вызваны его влиянием.
Изменения обилия, вызванные воздействием всех остальных факторов, необходимо по
возможности исключить. Для этого каждый раз рассматривалась лишь та часть
выборки, в которой были значимы различия только анализируемого фактора, а
проявление всех остальных учтенных варьировало несущественно.
Так,
при выявлении f(ain)
рассматривались изменения Nint внутри каждого
ключевого участка отдельно, т.е. по данным за один год наблюдений. Тем самым в
значительной степени исключалось влияние на неоднородность обилия следующих
факторов: bin, межгодовых
флуктуации численности популяций, различий широтно-климатических f(cni ) и долготных f(dni )
показателей территориальных выделов (координаты трансектов внутри каждого
ключевого участка различаются не более чем на 5').
Для
выявления f(bin) сравнивались исходные значения обилия в выделах (Nin
emp). При этом в анализе использовались
лишь данные с тех трансектов, на которых количество предпочитаемого видом i субстрата не выходло за пределы
интервала 0,5 – 0,7. Как и в случае с предыдущим фактором изменения, связанные
с bin рассматривались для каждого
ключевого участка отдельно.
F(cni) и f(dni) выявлялись
в процессе сравнения показателей реальной (имманентной) плотности вида (Nint).
При этом сопоставлялись средние арифметические для каждого ключевого участка
значения Nint (т.е. исключалось влияние bin). С каждого ключевого участка
по каждому виду i для
расчетов таких средних использовались данные из тех выделов, которые близки по
количеству пригодного для вида i субстрата
(для большинства видов ain=0,5 –
0,7). Влияние dni на широтно-климатические
изменения обилия исключалось тем, что каждый из участвующих в расчете
широтно-климатических рядов составлен данными, полученными в пределах
долготного сектора предпочтения вида i. В свою очередь, долготные ряды анализировались только по
данным, полученным в широтно-климатической зоне предпочтения этого вида. В
последнем случае различия широтных координат ограничены еще и 4º. Следует
признать, что при анализе изменений обилия по этим двум градиентам исключить
влияние межгодовых флуктуаций численности не удалось.
Как
уже упоминалось, на современном уровне изученности наиболее значимые для
большинства видов сочетания климатических факторов не определены. В настоящей
работе для характеристики широтных изменений обилия рассчитаны коэффициенты регрессии
отдельно по двум характеристикам среды: географическая широта места проведения
учета птиц (как маркер климатических параметров) и продолжительность
бесснежного периода с положительными среднесуточными температурами. Эти
климатические сведения заимствованы из Климатического атласа СССР [1960],
следовательно, данная величина – средняя многолетняя, а не та, которая была в
год исследований.
Последнее
сочетание двух климатических параметров скорее всего
не значимо непосредственно для большинства видов. Выбор его в качестве маркера
значимых для каждого вида, но пока еще не выявленных климатических условий
обусловлен следующим. Продолжительность безморозного периода выбрана потому,
что, во-первых, с ней уже отмечалась связь обилия птиц [Пузаченко, 1967].
Во-вторых, эта характеристика на равнине в той или иной мере скоррелирована с
продолжительностью периода с более высокими температурами. А, как известно, размножение
каждого вида наиболее продуктивно только при определенных температурах в определенный
период времени [Юрлов и др., 1994]. Отсутствие снежного покрова, особенно при
положительных температурах, создает благоприятные условия для выкармливания
птенцов большинства видов (вегетация растительности, активность беспозвоночных,
возможность добывать корм на почве и в воде и т.д.). В перспективе еще
предстоит выявить для каждого вида интегральную характеристику, которая наиболее
корректно отображает климатическую составляющую его экологической ниши.
Все
величины, являющиеся функциями по факторам среды, представлены нормированными
коэффициентами, выраженными в долях от единицы. Характеристики обилия (Ni), в том числе и идеальная
плотность видов, приводятся в количестве особей на 1 км2,
масса птицы – в граммах, количество субстрата и доля площади, занятая определенной
топоархитектурой в выделе, – в долях от единицы. Географические координаты
ключевых участков определены с точностью ±5'. Расстояния от территориальных
выделов до широтно-климатических зон и долготных секторов предпочтения
рассчитаны по физической карте (М 1: 2 000 000) и выражены в километрах.
Таким образом, результирующее модельное уравнение будет составлено из функций, полученных на весьма ограниченном наборе видов и иллюстрирующих идеальные представления. Следовательно, необходима оценка того, насколько совпадают моделируемые данным уравнением параметры с эмпирическими по всем изученным видам. Для этого на основе результирующего уравнения и сведений о географических и экологических предпочтениях видов, приведенных в классификационной таблице (см. табл. 1), выполнен прогноз обилия 186 видов птиц на 164 обследованных территориальных выделах. Прогнозные значения сопоставлены с эмпирическими. Для оценки использованы все имеющиеся в распоряжении материалы. Для видов, которым свойственна сезонная смена субстрата, в классификационной таблице указан не один, а два предпочитаемых типа субстрата. Для таких птиц при прогнозе в уравнение подставлялись средние арифметические количества субстратов обоих типов.
В
модели использованы зависимости обилия птиц от факторов в гнездовой период. В
то же время для многих, в первую очередь редких, птиц установить сроки
размножения в годы исследований не удалось. Поэтому все имеющиеся контрольные
эмпирические значения обилия усреднены за весь период наблюдений (первую половину
лета), который для большинства видов включает в себя не только гнездовой
период, но и время миграций или вылета молодых. Для приближения прогнозных
показателей к эмпирическим в расчетах дополнительно использован нормированный
коэффициент (kmg), характеризующий влияние миграций и вылета молодых на обилие. Значения его подобраны на основе
идеализированных представлений об изменениях количества мигрантов в зависимости
от географического положения территории [Юдкин, 1986а]. Кроме того, при
расчетах использован нормированный коэффициент, характеризующий влияние
антропогенного беспокойства и прямого преследования птиц (kant). Поправка
на это влияние вносилась для обилия наземногнездящихся видов, как наиболее уязвимых в гнездовой период к присутствию большого количества
людей и домашних животных. Ее величина весьма приблизительная, так как рассчитана
по небольшому количеству данных, заимствованных из некоторых публикаций
[Равкин, 1973, 1978; Равкин, Лукьянова, 1976; Юдкин, 1986б] Эта поправка
вносилась лишь для обилия в населенных пунктах и их ближайших окрестностях.
Так, для видов массой менее 35 г в населенных пунктах принят kant=0,1, массой более 35 г kant=0,
а в их окрестностях в радиусе 1 км kant=0,3. Во всех остальных случаях влиянием антропогенного
беспокойства пренебрегаем, т.е. kant=1. Оба эти
коэффициента в уравнении поставлены вместо k1,…,z, т.е. k1,...,z=kmg kant.
В
предлагаемой модели совокупность функций по факторам моделирует только характер
территориальной неоднородности обилия, независимо от его абсолютного уровня для
каждого вида. Понятие “характер неоднородности обилия” дано в
гл. 2. Абсолютный уровень обилия, на фоне которого формируется неоднородность,
согласно данной концепции определяется с учетом идеальной плотности населения
вида. Следовательно, оцениваться будут вначале эффективность только факторной
составляющей, а уже потом работа всей модели в целом, т.е. с добавлением
моделирования идеальной плотности. На каждом из этапов использована своя мера
сходства. Так, мерой сходства характеров территориальной неоднородности
прогнозного и эмпирического обилия каждого вида и суммарной плотности населения
всех видов послужил коэффициент их корреляции. Сходство абсолютных значений
модельного обилия с соответствующими им эмпирическими
оценивалось по доле учтенной прогнозом дисперсии эмпирических значений (Dσ 2). Эта доля
для каждого вида и для населения всех птиц в целом рассчитана следующим
образом:
Dσ 2=(σ 2emp- σ 2r) / σ
2emp,
где σ 2r – дисперсия значений, которые
являются разностью эмпирических показателей обилия и соответствующих им
прогнозных; σ2emp –
дисперсия эмпирических значений обилия.
3.2. Уравнения регрессии обилия по отдельным факторам
3.2.1. Величина птиц
За
характеристику величины птиц в данном случае принимается масса их тела. Зависимость
идеальной плотности населения видов от средней массы одной их особи имеет
довольно отчетливый гиперболический характер (рис. 11, а). Наиболее удачно эти изменения аппроксимируются функцией
lnNi0= -0,78·ln(0,0001mi)
или (4)
Ni0=(0,0001mi)-0.78
где mi – средняя масса одной особи
вида i
(рис. 11, б).
Рис. 11. Представление о
зависимости идеальной плотности населения вида (Ni0)
от массы одной его особи (m).
а – эмпирические
данные; б – идеальная кривая Ni0
(такой величины должна достигать Nint тех видов, численность
которых существенно не лимитируется емкостью мест зимовок; реальная
(имманентная) плотность Nint остальных видов тем
ниже Ni0,
чем меньше емкость мест их зимовок относительно емкости гнездовых территорий;
значения их обилия на графике должны располагаться в пределах заштрихованной
площади).
Следует
отметить, что суммарная биомасса всех птиц, составляющих реальную (имманентную)
плотность каждого вида (Nint), согласно
имеющимся данным возрастает с увеличением массы одной особи, особенно интенсивно
для мелких видов (см. табл.2). Таким образом, при идеальной плотности вида i суммарная биомасса всех составляющих
ее птиц (Mi0) зависит
от средней массы одной особи вида приблизительно следующим образом: Mi0=104 (0,0001mi)0.22. Вполне возможно, что суммарная энергия, трансформируемая количеством
птиц Ni0, – величина
для всех видов постоянная, независимая от их размеров. Во всяком случае эта энергетическая характеристика, рассчитанная по
формулам зависимости метаболизма от массы тела [Гаврилов, 1977] для количества
птиц Nint , оказалась
для мелких воробьиных несколько выше, чем для всех остальных видов. Но внутри
последней совокупности для этой величины не прослеживается каких-либо изменений,
совпадающих с изменениями массы тела одной особи. Большинство значений попадает
в интервал 0,5 – 2,0 тыс. ккал/(сут км2)
(см. табл. 2). Можно предположить, что в одинаково благоприятных ситуациях
одинакова и интенсивность трансформирования энергии населением разных видов,
независимо от величины их особей.
3.2.2. Топоархитектура
Обилие
птиц в зависимости от доли площади bi территориального
выдела n, на которой представлена предпочитаемая данным видом
топоархитектура, изменяется линейно (рис. 12, а). Такая зависимость вполне предсказуема, если исходить из
утверждений, приведенных в гл. 2. Эта линейность
прослеживается тем отчетливее, чем надежнее имеющиеся показатели обилия (т.е.
вид более заметный, а его численность выше). Следовательно
f(bin)=bin, и
показатель обилия может быть рассчитан по формуле
Nin=Ni0·bin·kf , (5)
где kf – произведение функций по
другим факторам. Если в уравнение (5) на место bin подставить
не долю площади, выраженную в долях от единицы, а абсолютную площадь какой-либо
территории, то на месте Nin будет уже не обилие
(относительный показатель на единицу площади), а численность, т.е. абсолютное
число птиц на этой площади.
Рис. 12. Примеры зависимости обилия вида (Ni) от доли площади с
предпочитаемой видом топоархитектурой bi (а) и от количества пригодного субстрата ai (б).
1 – Anthus trivialis (в среднем за гнездовой период 1982 г., 57°00'
с.ш., 66°45'в.д.);
2 – Sylvia borin (там же); 3
– Parus montanus (в среднем за первую
половину лета 1981 г., 60°25'
с.ш., 69°50'
в.д.); 4 – Emberiza aureola (там же); 5 – Anthus
trivialis (в среднем за гнездовой период 1992 г., 58°10'
с.ш., 65°25'
в.д., там же и все последующие виды); 6 – Sylvia
borin; 7 – Acrocephalus dumetorum;
8 – Motacilla flava.
3.2.3. Субстрат
Характер изменения обилия вида в зависимости от количества соответствующего субстрата (ain) просматривается не столь отчетливо (см. рис. 12, б). Однако для большинства видов можно констатировать нечто общее. Так, за редким исключением, большинство видов при прочих благоприятных условиях встречалось на трансекте только тогда, когда там имелся соответствующий субстрат. При ain менее 0,5 подавляющее большинство показателей обилия можно охарактеризовать как низкое для этого вида (ниже среднего по выборке). Причем в интервале 0 – 0,5 прослеживается тенденция к увеличению числа птиц с ростом количества субстрата. При значениях последнего 0,5 – 0,7 большинство показателей обилия можно отнести к разряду низких или средних. В указанном интервале направленные изменения обилия не прослеживаются. Для тех территорий, на которых ain превышает 0,7, характерно в основном большое количество птиц, и оно, судя по всему, на этом отрезке оси возрастает экспоненциально.
Отсутствие
видимых направленных изменений обилия в интервале количества субстрата 0,5 –
0,7 можно объяснить двумя причинами. Возможно, что такое ain наиболее часто встречается
в природных экосистемах. Так, в имеющейся выборке оно присутствует более чем в
трети ненулевых значений. Не исключено, что поведение большинства видов
приспособлено в наибольшей степени к этому интервалу, внутри которого для птиц
изменения незначимы, а выход за его пределы весьма ощутим. Но возможно и то,
что именно в этом интервале исследователь меньше всего улавливает различия в
количестве субстрата, а потому и оценивает его наименее адекватно.
Приняв
любое из объяснений, целесообразно аппроксимировать изменения обилия так, чтобы
нормированный коэффициент, полученный при расчете f(ain ), был равен единице в том случае, когда
ain попадает в
середину интервала 0,5 – 0,7. По тем же соображениям эталонное количество
субстрата для идеальной плотности вида принято равным 0,6. Для отображения
такой зависимости оптимальным будет выражение
f(ain)=exp[20(ain-0.6)3]
или
(6)
lnf(ain)= 20(ain-0.6)3.
Согласно
концептуальному уравнению (3), зависимость обилия от совокупного влияния
количества топоархитектуры и субстрата должно быть выражено f(bin ) f(ain
)=bin exp[20(ain-0,6)3]. Графическое отображение
зависимости обилия вида одновременно от двух этих факторов представлено на рис.
13. Такой характер изменений присущ для любой ограниченной территории, внутри
которой широтными и долготными различиями можно пренебречь, а все остальные
значимые для вида условия сходны.

Рис. 13. Зависимость обилия вида (Ni) от совокупного
воздействия доли площади с предпочитаемой видом топоархитектурой (bi) и количества
пригодного субстрата (ai).
3.2.4. Широтно-климатические и долготные факторы
Изменения
обилия, связанные с отклонением как от широтно-климатической зоны, так и от
долготного сектора предпочтения точнее всего иллюстрируются S-образной кривой (рис. 14). Вполне
логично, что эта линия близка к половине классической кривой толерантности.
Представленные изменения можно аппроксимировать одной функцией, но с разными
коэффициентами регрессии:
f(cni )=exp[-(xci·cni )rci]
или lnf(cni )= -(xci·cni )rci (7)
f(dni )=exp[-(xdi·dni )rdi]
или
lnf(dni)= -(xdi·dni )rdi (8)
где
коэффициенты xci
и xdi и
показатели степени rci и rdi принимают
специфические для вида i значения в уравнениях, характеризующих
изменение обилия по широтно-климатическому градиенту (7) и по долготе (8) соответственно.
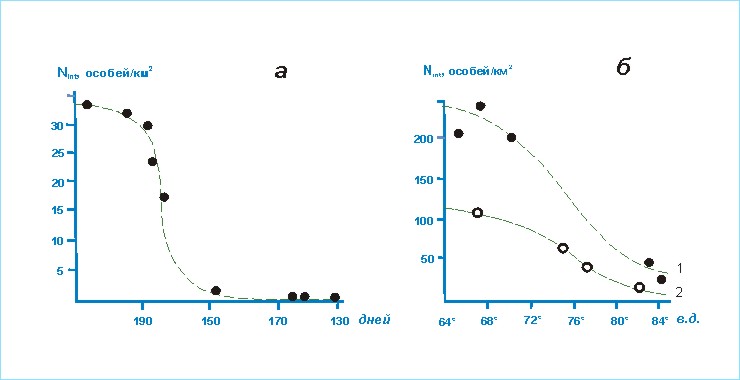
Рис. 14. Изменение обилия
вида (на примере Nint):
а
– Sitta europaea при удалении от
широтно-климатической зоны предпочтения к северу (северная граница этой зоны 55°
с.ш.); б – те же данные по плотности
вида, но по оси абсцисс – продолжительность бесснежного периода с положительными
среднесуточными температурами; в – Phylloscopus trochilus при отклонении к
востоку от долготного сектора предпочтения (восточная граница этого сектора
проведена по меридиану 60°
в.д.); 1 – в
пределах полосы 57 – 61°
с.ш.; 2 – 63 – 66°
с.ш.
Для уравнения (7) коэффициенты xci и rci
оптимизировались отдельно для случая, когда cni выражено в расстоянии от широтной зоны предпочтения,
и отдельно для случая, когда cni выражено в
продолжительности бесснежного периода при положительных среднесуточных
температурах (табл.3). При этом оказалось, что внутри каждой
широтно-климатической группы птиц коэффициенты регрессии различаются очень незначительно.
Более того, вариабельность значений как самих
коэффициентов, так и границ зон предпочтения существенно меньше в том случае,
когда широтно-климатический фактор выражен не линейным расстоянием, а сочетанием
климатических характеристик.
Т а б л и ц а 3
Коэффициенты регрессии изменения обилия перелетных птиц по фактору
отклонения от их широтно-климатической зоны предпочтения
(для уравнений (4), (6), (7))
|
|
Среднее
значение маркера фактора, при котором начинается уменьшение реальной плотности
видов |
Средний
коэффициент регрессии по фактору отклонения, выраженному |
|
||||
|
Широтно-климатическая
группа птиц |
Географическая
широта в секторе 65-77º в.д., град. с.ш. (M±m) |
Продолжительность
бесснежного периода с положительными среднесуточными температурами, дней (M±m) |
линейным расстоянием,
км |
продолжительностью
бесснежного периода с положительными среднесуточными температурами, дней |
n |
||
|
|
|
|
xci (M±m) |
rci |
xci (M±m) |
rci |
|
|
При удалении от
южной границы: |
|
|
|
|
|
|
|
|
гемиарктическая |
70,5±0,4 |
108±0 |
16·10-4±9·10-5 |
2 |
0,032±0,002 |
3 |
2 |
|
гипоарктическая |
66±0,3 |
132±0,2 |
29·10-4±27·10-5 |
2 |
0,045±0,003 |
3 |
9 |
|
бореальная |
62±0,4 |
161±0,3 |
32·10-4±18·10-5 |
3 |
0,082±0,008 |
3 |
4 |
|
При удалении от
северной границы: |
|
|
|
|
|
|
|
|
бореальная |
66±0,4 |
132±0,4 |
52·10-4±63·10-5 |
3 |
0,09±0,009 |
3 |
4 |
|
трансумеренная |
63 |
160 |
15·10-4 |
3 |
0,02 |
3 |
1 |
|
северная умеренная |
62±0,7 |
162±2,4 |
21·10-4±18·10-5 |
2 |
0,037±0,006 |
3 |
3 |
|
среднеумеренная |
58±0,2 |
178±0,4 |
25·10-4±18·10-5 |
3 |
0,058±0,003 |
3 |
12 |
|
южная умеренная |
55±0,2 |
185±0,4 |
20·10-4±9·10-5 |
3 |
0,061±0,002 |
3 |
10 |
|
остальные более “южные”виды |
53±0,4 |
187±1,3 |
23·10-4±18·10-5 |
4 |
0,13±0,012 |
3 |
5 |
Для изменений по долготе (8) значения коэффициентов xdi и rdi удалось рассчитать лишь для
видов двух долготных групп – срединно-западной и срединно-восточной. Для птиц,
вошедших в первую группу, при отклонении от предпочитаемого сектора к востоку xdi =1,2·10-3,
а для видов, составивших вторую группу, при отклонении к западу xdi =0,9·10-3.
В обоих случаях rdi
=2. Эти значения коэффициентов получены лишь в пределах средне-
и южно-умеренной широтно-климатических зон.
Как
видно из табл. 3, уменьшение
обилия по климатическому градиенту в большинстве случаев несколько интенсивнее
при отклонении от зоны предпочтения к северу, чем к югу. Это заметно и для
одного вида, и для разных видов в тех случаях, когда первое явление у одних видов наблюдается на тех же широтах, что второе у
других. Объяснить подобное можно с помощью гипотезы, которая используется для
идеальной ситуации в условиях равнины. Эта гипотеза принимается при следующих
допущениях: климатические условия меняются только в широтном направлении, а все
прочие условия повсюду равны. Тогда линия кратчайшего расстояния от зимовок до
широтно-климатической зоны предпочтения совпадает с меридианом.
Суть
данной гипотезы заключается в следующем. Согласно обсуждаемой концепции, самые
благоприятные для размножения популяции условия складываются не во всей широтно-климатической
зоне предпочтения и только на тех долготах, на которых находятся места ее
зимовок (в точке p3, рис. 15).
При перелетах с зимовок в эти места расстояние, которое вынуждены преодолевать
птицы, будет минимальным (op3). Птицы,
летящие в точку p3 , имеют
преимущества перед птицами, которые будут лететь правее или левее данной точки
в пределах благоприятной широты. Это преимущество заключается в необходимости
тратить меньше времени и энергии на подобный перелет.
Рис. 15. Схема
пространственных изменений совокупного воздействия на птиц
широтно-климатического и долготного факторов (обозначения в тексте).
Расселение
популяции с максимально благоприятной территории (p3) в любую
сторону от широтно-климатической зоны предпочтения сдерживается ухудшением для
нее климатических условий. Негативное воздействие последних
возрастает по мере удаления от зоны предпочтения. При этом расселение популяции
из благоприятной зоны строго в направлении зимовок будет сдерживаться только
одним климатическим фактором. Расстояние необходимых миграционных перелетов при
этом будет уменьшаться (op5 < op3). Расселение
в противоположную от зимовок сторону будет сдерживаться уже двумя факторами. В
таком случае негативное влияние климатических факторов усугубляется
необходимостью совершать птицами более длительные перелеты, так как op1 > op3. Ограничивающее
влияние «лишнего пути» на расселяющихся птиц заключается в том, что особи, миграционный
путь которых короче, имеют перед ними преимущества (при прочих равных затрачивают
меньше времени и энергии на перелет).
Если
придерживаться принятой гипотезы, то расселение в пределах благоприятной
широтно-климатической зоны в сторону от долготы предпочтения будет сдерживаться
лишь одним фактором – необходимостью совершать более длинный перелет (op4 > op3). Причем
значимость этого фактора «лишнего расстояния» для разных птиц будет различной.
Известно, что физиологические показатели, в первую очередь особенности метаболизма,
у видов (или популяций), приспособленных к перелетам, существенно отличаются от
этих показателей у оседлых птиц [Гаврилов, Дольник, 1976; Гаврилов, 1995].
Таким образом, первые виды не могут переносить низкие отрицательные температуры,
а вторые – совершать дальние перелеты. Исходя из этого вполне
логично предположить: чем к более длительным перелетам адаптирован вид,
тем меньшее сдерживающее влияние на его распространение будет оказывать фактор
«лишнего расстояния». Скорее всего поэтому обилие
видов, приспособленных к дальним перелетам, по градиенту долготы изменяется с гораздо
меньшей интенсивностью, чем при отклонении от благоприятной широтно-климатической
зоны. В то же время для ближних мигрантов, и особенно для оседлых видов, фактор
«лишнего расстояния» более значим. Их обилие по широте и долготе изменяется с
более сходной интенсивностью.
На
основе изложенной гипотезы в концептуальную модель внесено дополнение,
относящееся к описанию некоторых изменений при удалении от зон предпочтения по
широтно-климатическому градиенту и долготе одновременно. Так, при удалении по
широте в сторону зимовок, но при малом угловом отклонении по долготе до определенного
его предела, расстояние op6,
преодолеваемое птицами, еще не будет превышать “норму” op3 (см. рис.
15). При таких отклонениях до определенного предела необходимость преодолевать
лишнее расстояние еще отсутствует, а ограничивающим обилие фактором будет лишь
широтно-климатический. По мере увеличения угла долготного отклонения при одном
и том же удалении по широте с момента, когда требуемый путь превысит «норму» op3, начнет
действовать фактор «лишнего расстояния», проявление которого будет нарастать (op8>op7>op3). Иными
словами, в данной ситуации вес фактора «лишнего расстояния» в совокупном
воздействии будет интенсивно увеличиваться. Для отображения изложенного
фрагмент f(cni
) f(dni
) концептуальной модели
целесообразно заменить одним выражением: f(cni,dni )=exp[-(xsi·hni )rsi]. В нем
значения коэффициентов xsi и rsi будут средними взвешенными
соответствующих коэффициентов из уравнений (6) и (7) (при cni , выраженном
в линейном расстоянии). Так, xsi=(xci·cni+xdi·dni
)/(cni+dni ), rsi=(rci·cni+rdi·dni )/(cni+dni ), а кратчайшее
расстояние отклонения от границ широтной зоны и долготного сектора предпочтения
(op3,…,8) запишется
как
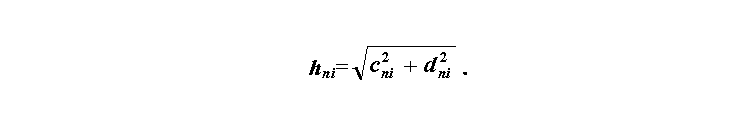
При
таком отображении анализируемых зависимостей в случаях с малыми отклонениями по
долготе, когда фактор «лишнего расстояния» еще не действует, соответственно и
вес коэффициентов из f(dni) в этой
средней будет мал. С увеличением отклонения по долготе по мере нарастания
влияния фактора «лишнего расстояния» будет возрастать и вес коэффициентов из f(dni), т.е. как
раз тех коэффициентов, которые характеризуют влияние фактора «лишнего расстояния».
Таким образом, функция изменения обилия при одновременном удалении по широте в
сторону зимовок и отклонении по долготе должна иметь вид
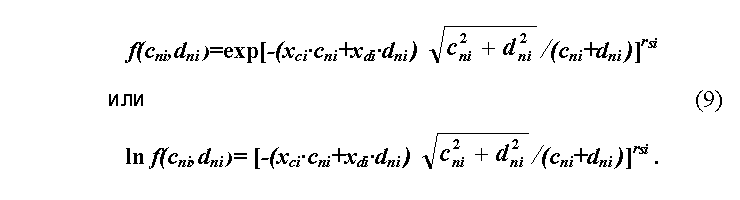
При
одновременном удалении по широте в противоположную от зимовок сторону и
отклонении по долготе на птиц воздействуют оба упомянутых фактора. По мере
отклонения по долготе вес фактора «лишнего расстояния» в совокупном влиянии
увеличивается гораздо меньше, чем при удалении в сторону зимовок, так как при
отклонении на одно и то же расстояние l всегда op8 /op5>>op2 /op1. В случаях удаления по широте в противоположную от зимовок сторону
лучше всего изменения обилия одновременно по двум факторам будет
отражать, как и предлагалось изначально, произведение функций (7) и (8). Это
произведение после преобразований примет вид
f(cni,dni )=exp{-[(xci·cni
)rci+(xdi·dni )rdi]}
или
(10)
ln f(cni,dni )= -[(xci·cni )rci +(xdi·dni ) rdi ] .
На рис. 16 показаны изменения обилия в умеренном поясе, рассчитанного по уравнению (9) для территории, расположенной южнее широтной зоны предпочтения, и по уравнению (10) для площади севернее этой зоны. В первом случае при отклонении, выраженном в линейном расстоянии, xci=20·10-4, rci=2, а при отклонении, выраженном в значениях климатического фактора, xci=0,035, rci=3. Во втором случае использованы коэффициенты: xci = 31·10-4, rci = 2 и соответственно xci = 0,08 и rci =3. Таким образом, рис. 16 иллюстрирует идеальную конфигурацию гнездового ареала перелетного вида на равнине, а также широтные и долготные изменения обилия внутри ареала. Естественно, что данная схема может рассматриваться лишь при следующих допущениях: климатические условия на всей площади изменяются только по градиенту широты, а изменения ускоряются в северном направлении, все остальные условия повсюду равны и благоприятны для вида.
Рис. 16. Идеальные представления об изменениях реальной (имманентной)
плотности перелетных видов в условиях равнин по градиентам
широтно-климатической совокупности факторов и географической долготы.
По оси абсцисс –
расстояние от широтного сектора предпочтения, по оси ординат – маркер
широтно-климатических факторов; Сплошные и штриховая
замкнутые изолинии соединяют точки с одинаковыми значениями реальной плотности
вида i, 0,01–1 – значения
нормированных коэффициентов реальной плотности (Ñint); горизонтальные
изолинии соединяют точки с одинаковыми значениями продолжительности бесснежного
периода при положительных среднесуточных температурах, z,.., z-30, t,..,
t+40 – количество дней,
z – минимальное
значение предпочитаемого видом i интервала,
t – максимальное; wº и y° – значение меридианов, являющихся
соответственно западной и восточной границами долготного сектора предпочтения
вида i; насыщенность заливки пропорциональна величине плотности;
А
– район зимовок вида i.
Нетрудно убедиться, что ареалы многих видов, в основном те, которые простираются на равнине и не слишком велики по площади, своими очертаниями близки к изображенному на рис. 16. В то же время предпочитаемые на равнине видом i климатические условия формируются и в горах (в северном полушарии только в тех, которые расположены южнее, чем зона широтно-климатического предпочтения на равнине). Следовательно, изменения обилия равнинных видов в горных странах, вызванные совокупным проявлением долготного и климатического факторов, должны описываться уравнением (9). При этом для тех горных районов, которые расположены не далее op3 от мест зимовок (см. рис. 15), подобный расчет целесообразно упростить до уравнения (8).
Итак,
если принять предлагаемую концепцию, то из приведенных рассуждений вытекает
следующее. Не только для оседлых, но и для кочующих и перелетных видов величина
и местоположение гнездовых ареалов, помимо пригодных для размножения условий, определяются
соответственно доступной емкостью и локализацией мест, где возможно переживание
самого неблагоприятного периода – зимы. Несомненно, что в этот период дефицит
пищи испытывают не только птицы, которые зимуют в холодном климате, но зачастую
и дальние мигранты, проводящие зиму в низких широтах. Наиболее вероятно, что последние вынуждены вступать в трофическую конкуренцию с
другими зимующими совместно с ними мигрантами и с более специализированными
аборигенами. Возможно, поэтому у большого количества видов при кажущейся
обширности пригодных для зимовки площадей зимовочные ареалы гораздо меньше, чем
гнездовые. Вероятно, только в таких местах более сильные конкуренты по
каким-либо причинам не могут реализовать свое преимущество перед этими видами.
Результирующее
уравнение регрессии максимально возможного гнездового обилия птиц на любой
территории принимает следующий вид:
Nin=(0,0001mi )-0.78 bin exp[20(ain-0,6)3] w k1,...,z
или (11)
ln Nin=
-0,78 ln (0,0001mi )+ln bin+20(ain-0,6)3+w+ln k1,…,z .
Здесь
w принимает
значение правой части уравнения (9) или (10), в зависимости от того, к северу
или к югу от широтной зоны предпочтения расположен анализируемый участок. В
мелком масштабе общая тенденция этих изменений будет близка к
изображенным на рис. 16, а в крупном – в первую очередь будут превалировать
территориальные изменения, показанные на рис. 13.
Эта
модель гносеологическая, т.е. претендует главным образом на описание принципиального
характера территориальных различий обилия птиц и основных причин, вызывающих
их. Полученные коэффициенты регрессии весьма приблизительны. Не исключена
существенная их корректировка после специальных полевых исследований.
Не
следует забывать, что предлагаемая модель создана только для репродуктивного
периода. Вне этого периода понятие Ni0 теряет свою
актуальность, поскольку птицы уже не привязаны к определенной территории. При
быстром использовании всего доступного кормового ресурса на ограниченной
площади им ничто не мешает покинуть эту территорию. Для такой утилизации
ресурса в послегнездовой период птицы, как правило, концентрируются в кормных
местах с плотностью, значительно превышающей Ni0. Кроме
того, после гнездования почти у всех видов связь с топоархитектурой, как с
фактором защищенности, должна ослабевать, а с субстратом, который отчасти
характеризует трофический фактор, оставаться сильной. Следовательно, для
послегнездового времени в уравнение регрессии необходимо вносить существенные поправки.
3.3. Степень соответствия моделируемых параметров эмпирическим данным
Прогноз
характера территориальной неоднородности обилия отдельных видов выполнен на
основе уравнения (11). Для всех видов средняя
арифметическая коэффициентов корреляции модельных и эмпирических значений составила
0,45±0,2, а их средняя взвешенная (с
учетом обилия видов) 0,69±0,03.
Коэффициенты корреляции для отдельных видов распределяются следующим образом.
Для 27 видов (15% всех анализируемых) они попадают в интервал от –0,02 до
+0,14, т.е. ниже критического значения (при 5%-м уровне значимости). За
исключением двух видов, это птицы или чрезвычайно редкие на обследованной
территории, или плохо заметные для учетчиков. В обоих случаях имеющаяся протяженность
учетных маршрутов слишком мала для достоверной оценки их обилия. Имеющиеся эмпирические
данные по таким птицам слишком ненадежны и не могут служить хорошим контролем.
Для 22 видов (12%) получена слабая положительная корреляция, превышающая
критическое значение (0,15-0,25). За исключением двух видов это также или
редкие птицы, или обычные, но плохо заметные. Более сильная положительная
корреляция (0,26 – 0,50) обнаружена для 63 видов (34%). Больше половины их
широко распространены и обычны на изучаемой территории, но особенности их
поведения также не дают возможности достаточно надежно оценить их обилие при
выбранной норме учета. Следовательно для них нет
смысла ожидать более сильной сопряженности прогнозных и эмпирических значений.
Другая часть этой группы – хорошо заметные обычные и многочисленные виды. Наибольшие
коэффициенты корреляции (0,51 – 0,99) обнаружены для 71 вида (39%). За редким исключением
это хорошо заметные обычные или многочисленные виды, для оценки
обилия которых использованная протяженность учетов достаточна.
Абсолютные
значения обилия рассматриваемых видов смоделированы менее удачно, чем характер
его неоднородности. Так, простая средняя значений доли
учтенной прогнозом дисперсии показателей обилия составила всего лишь 9±1%, а
средняя взвешенная 30±4%.
Таким
образом, характер пространственной неоднородности обилия сравнительно успешно
спрогнозирован для большего числа видов, чем абсолютные значения обилия. Такое
расхождение произошло за счет того, что для многих видов показатели модельного
обилия оказались повсеместно выше соответствующих им эмпирических. Почти для каждого
из видов это превышение примерно одинаково во всей выборке. Такое систематическое
завышение вполне объяснимо, поскольку абсолютный уровень обилия в данной модели
рассчитан на основе идеальной плотности вида. В то же время, как уже
упоминалось, если численность вида лимитируется каким-либо ресурсом на
зимовках, то его Nint нигде не должно достигать уровня Ni0 , а в
данных расчетах лимитирующее влияние зимовок не учтено (как и ряда других
факторов). При этом вполне очевидно, что для таких видов на имеющихся
материалах легко подобрать в данную модель дополнительные коэффициенты, которые
существенно улучшат совпадение прогнозных абсолютных значений обилия с эмпирическими. Однако сами коэффициенты и их величины не
будут представлять познавательной ценности, пока у них не появится предметное
толкование. В настоящей работе эти поправки не рассчитывались, потому что выполненный
прогноз не является целью. Он – всего лишь инструмент для оценки того, насколько
идеальные представления о зависимостях далеки от самих зависимостей, которые характерны
для выборки эмпирических данных.
Поскольку прогноз выполнен почти для всех видов, встреченных
в обследованных выделах, в результате получились модельные
орнитокомплексы этих выделов. Появилась возможность оценить их сходство с
контролем. Так, неоднородность суммарной плотности населения птиц, обитающих в
каждом выделе, спрогнозировано с такой же полнотой, как неоднородность обилия
большинства видов. Коэффициент корреляции модельных и эмпирических значений
суммарной плотности составил 0,88. Абсолютные значения смоделированного
суммарного обилия птиц в каждом выделе меньше совпадают с соответствующими
эмпирическими: прогнозом учтено лишь 52% дисперсии этих показателей. Причем
показатели модельной плотности населения всех птиц опять же выше соответствующих
им эмпирических (за счет упомянутого систематического завышения прогнозного
обилия ряда видов).
Успешность прогнозирования характера территориальной
неоднородности облика населения (видового состава и обилия каждого вида)
оценивалась по корреляция
двух матриц коэффициентов Жаккара – Наумова [Наумов, 1964]. Одна из этих матриц
содержит коэффициенты, которые рассчитаны между всеми модельными орнитокомплексами,
а другая состоит из коэффициентов между эмпирическими вариантами населения.
Коэффициент корреляции между двумя матрицами составил 0,78. Если исключить
влияние межгодовых флуктуаций численности, т.е. рассматривать такую сопряженность
лишь по одному году наблюдений, то средняя коэффициентов корреляции для 11 пар
матриц (каждая по отдельному ключевому участку) оказалась еще выше (0,86±0,02).
В то же время попарное сходство каждого из модельных
орнитокомплексов с соответствующим ему контрольным для большинства вариантов
невелико, средняя арифметическая таких коэффициентов Жаккара – Наумова
составила 27±1%. Во
многом столь низкие значения коэффициента являются результатом систематических
завышений прогнозного обилия ряда видов. Кроме того, не следует
упускать из вида особенности самого коэффициента, основная из которых – нелинейность
его изменений. Для иллюстрации укажем, что значения этого коэффициента,
характеризующие сходство результатов учетов двух исследователей, идущих
одновременно по одному и тому же маршруту друг за другом, зачастую бывают не
слишком высокими. Так, на открытых пространствах в подобных случаях коэффициенты
сходства варьируют от 40 до 60%. Лишь в самый разгар гнездования и в основном
на лесных территориях, где большинство птиц учитывается по пению территориальных
самцов, результаты синхронных учетов двух специалистов совпадают больше
(60-90%).
Итак,
удачным можно считать прогноз характера территориальной неоднородности обилия
птиц. Иными словами, достаточно хорошо работает та часть модели, которая характеризует
зависимость обилия от значимых факторов в районах гнездования. Для более
успешного моделирования абсолютного уровня обилия необходима еще информация об
экологической емкости зимовок вида. При этом неоднородность параметров,
характеризующих орнитокомплексы в целом, смоделирована удачнее, чем
территориальные отличия обилия большинства видов в отдельности. Последнее в
значительной степени обусловлено тем, что расчеты делались не по проявлениям
конкретных факторов, к которым адаптирован каждый вид. Вместо количественной
оценки каждого такого фактора в расчетах использована суммарная оценка всех
сходных факторов, отнесенных к одному типу.
ЗАКЛЮЧЕНИЕ
Предлагаемая концептуальная модель иллюстрирует сущность и характер пространственных изменений обилия под влиянием внешних факторов. Основу модели составляет описание влияния наиболее значимых из них. Таковыми считаются те факторы, проявление которых характеризуется наибольшей пространственной неоднородностью. Следовательно, что именно они на наибольшей площади выступают в качестве лимитирующих. В разное время и в различных местах может проявляться воздействие и множества других локальных или временных факторов, некоторые из них могут быть видоспецифичными. В результате реальный характер территориальных изменений обилия у разных видов в той или иной степени отличается от того идеального, который моделируется предлагаемым уравнением. Несмотря на эти искажения, все же не выявлено принципиальных различий между реальным и идеальным характерами территориальной неоднородности обилия большинства видов, во всяком случае по имеющимся материалам. В то же время в модели предусмотрена и продемонстрирована в расчетах возможность учесть влияния каких-либо дополнительных факторов (на примере фактора беспокойства).
Изложенные
представления об организации распределения птиц позволяют сделать ряд выводов
об изменениях обилия в пространстве и времени:
-
потенциальное обилие вида зависит от средних размеров тела одной его особи: чем
мельче вид, тем при одинаково благоприятных условиях он достигает большего
обилия;
-
обилие определенного вида на каком-либо участке определяется долей пространства,
на котором представлена экологическая ниша этого вида;
- обилие определяется также расстоянием от этого участка до места, где данный вид имеет возможность благополучно перезимовать. При прочих равных условиях, чем больше это расстояние, тем меньше обилие вида на участке.
Следует
иметь в виду, что по характеристикам выбранных участков можно судить лишь о
характере пространственной неоднородности обилия. Абсолютная же величина
обилия, помимо размеров вида и условий на участке, зависит еще от общей
численности популяции. При этом изменения общей численности вида или отдельной
популяции неоднозначно отражаются на его обилии на
отдельных участках. Сама численность может лимитироваться двумя совокупностями
факторов. Первая – это площадь пригодных (по всем биотическим и абиотическим
факторам) для успешного размножения мест. Причем значима не только их площадь в
современном ареале вида, но и за его пределами, особенно вблизи границ. Вторая
совокупность – это емкость зимовок. У тех видов, численность которых лимитируется
емкостью зимовок, в наиболее благоприятных условиях реальная (имманентная)
плотность должна быть ниже идеальной. Именно у таких видов гнездовые ареалы
будут заметно меньше, чем у сходных по размерам более
обильных видов. Последние, заняв с максимально возможным обилием все
пригодные площади на долготе зимовок, вынуждены гораздо шире расселяться по
градиенту долготы.
Общая
численность популяции в зависимости от ситуации может по-разному определять
обилие вида на каком-либо конкретном участке. Так, при большей общей численности
популяции выше будет и реальная (имманентная) плотность вида. Это значит, что
при прочих равных условиях обилие вида на конкретном участке будет также выше.
Если же в ареале и за его пределами пригодных площадей станет меньше, а общая
численность не изменится, то будет возрастать реальная (имманентная) плотность
вида (за счет повышения концентрации птиц на оставшихся пригодных площадях).
Это, в свою очередь, может неоднозначно отразиться на обилии на отдельном участке.
Так, если внутри этого участка пригодная площадь не изменится, то обилие на нем
возрастет. Но такая концентрация без последствий может возрастать лишь до определенного
предела, которым в идеальных условиях является идеальная плотность. Если вид не
будет иметь возможности дальше расширять свой ареал по долготе (из-за отсутствия
пригодных мест), то после достижения этого уровня численность уже будет
лимитироваться пригодной для размножения площадью.
При
расширении по каким-либо причинам зимовочного ареала вида при прочих неизменных
условиях должен расширяться и его гнездовой ареал. Соответственно при этом
будет увеличиваться и общая численность вида. В такой ситуации на участках,
расположенных в том долготном секторе предпочтения, который был до расширения
зимовочного ареала, при прочих неизменных условиях обилие не должно заметно
измениться. За пределами этого сектора обилие должно увеличиться в той стороне
ареала, куда расширились зимовочные площади.
Перечисленные
выводы могут быть весьма полезными при изучении или характеристике
пространственного распределения птиц, при изучении их численности и ее многолетней
динамики. В большинстве случаев для таких исследований исходной информацией
служат показатели обилия на весьма ограниченном наборе обследованных участков.
Объективная оценка численности по таким показателям – задача гораздо более
сложная, чем это может показаться без учета всех перечисленных положений.
СПИСОК ЛИТЕРАТУРЫ
Бобков Ю.В., Торопов К.В., Шор
Е.Л., Юдкин В.А. К
орнитофауне южной тайги Западно-Сибирской равнины // Материалы к
распространению птиц на Урале, в Приуралье и Западной Сибири. -Екатеринбург, 1997. -С. 14-21.
Брунов В.В. Опыт
анализа фаунистических групп птиц тайги Палеарктики // Бюл. МОИП. Отд. биол. -1978.
-Т. 83, вып. 5. -С. 5-15.
Брунов В.В. О
некоторых фаунистических группах птиц тайги Евразии // Современные проблемы
зоогеографии. -М.: Наука, 1980. -С. 217-254.
Брунов В.В. О
существовании южнотундрово-лесотундровой географо-генетической группы птиц //
Современные проблемы биогеографии. -М.: Изд-во МГУ,
1982. -С. 74-91.
Бурский О.В. Гнездовое
размещение воробьиных птиц в енисейской тайге как отражение экологических
особенностей видов // Фауна и экология птиц и млекопитающих Средней Сибири. -М.: Наука, 1987. -С. 108-142.
Бурский О.В. Опыт анализа годовых изменений численности птиц //
Экология. -1993. -N 3. -С. 27-29.
Вартапетов Л.Г. Птицы
таежных междуречий Западной Сибири. -Новосибирск: Наука.
Сиб. отд-ние, 1984. -242 с.
Вартапетов Л.Г., Юдкин В.А. Птицы
Притымской средней тайги // Материалы к распространению птиц на Урале, в
Приуралье и Западной Сибири. -Екатеринбург, 1999. -С.
69-71.
Владышевский Д.В. Значение
трофического фактора для птиц в различных экологических ситуациях // Экология
популяций лесных животных Сибири. -Новосибирск: Наука.
Сиб. отд-ние, 1974. -С. 119-165.
Владышевский Д.В. Экология
лесных птиц и зверей (кормодобывание и его биоценотическое значение). -Новосибирск: Наука. Сиб. отд-ние, 1980. -264с.
Гаврилов В.В.
Пространственная структура популяций и зависимость размеров индивидуальных
участков от массы тела и кормности местообитаний у куликов северо-востока
Якутии // Орнитология. -М.: Изд-во МГУ, 1988. -Вып.
23. -С. 52-62.
Гаврилов В.М.
Биоэнергетика миграций // Методы изучения миграций птиц: Материалы Всесоюз. шк.-семинара. -М., 1977. -С. 7-16.
Гаврилов В.М. Количественные взаимосвязи между потреблением пищи,
энергетическим метаболизмом, температурой среды и размерами в семействе Corvidae // Орнитология –М.: Изд-во МГУ, 1995. Вып. 26. –С. 165-173.
Гаврилов В.М., Дольник В.Р. Годовые циклы метаболизма, поведения и состава тела у
перелетных и оседлых подвидов зяблика и домового воробья // Орнитология. –М.: Изд-во МГУ, 1976. Вып. 12. -С. 178-199.
Гиляров М.С.
Соотношение размеров и численности почвенных животных // Докл. АН СССР. -1944.
-Т.43, №6. -С. 283-285.
Гуреев С.П. Кузнецкий
Алатау // Пространственно временная динамика животного населения (птицы и мелкие
млекопитающие). -Новосибирск: Наука. Сиб. отд-ние,
1985. -С. 88-115.
Дольник В.Р. Затраты времени и энергии на защиту территории у
птиц: связь с размерами тела и территории, потребностями особи и выводка и с
экономией энергии при выкармливании птенцов // Экология. -1993. -№2. -С. 26-38.
Иваницкий В.В.
Пространственно-ориентированное поведение птиц. // Орнитология. –М.: Изд-во МГУ, 1998. -Вып. 28. -С. 6-25.
Касыбеков Э.Ш. Птицы
восточной части Иссык-Кульской котловины (численность, распределение и пространственная
организация населения): Автореф. дис. ... канд. биол. наук. -Новосибирск,
1990. -22 с.
Кашкаров Д.Н. Основы
экологии животных. -Л.: Гос. учеб. изд-во Наркомпроса
РСФСР, 1944. -384 с.
Кищинский А.А.
Орнитофауна северо-востока Азии. -М.: Наука, 1988.
-289 с.
Климатический Атлас
СССР. -М.: ГУГК, 1960.
Курлавичус П. Биотопическое распределение птиц в агронасаждениях. -Вильнюс: Мокслас, 1986. -108 с.
Любищев А.А. О форме
естественной системы организмов // Проблемы формы систематики и эволюции организмов.
-М: Наука, 1982. -С. 24-36.
Малков Н.П., Равкин Ю.С.
Пространственная структура населения птиц. Центральный Алтай // Пространственно-временная
динамика животного населения (птицы и мелкие млекопитающие). -Новосибирск:
Наука. Сиб. отд-ние, 1985. -С. 115-131.
Наумов Р.Л. Птицы в
очагах клещевого энцефалита: Автореф. дис. ... канд. биол. наук. – М., 1964.
-19 с.
Познанин Л.П.
Экологические аспекты эволюции птиц. -М.: Наука, 1978.
-147 с.
Преображенская Е.С. Кормовое
поведение и биотопическое распределение воробьиных птиц Приветлужья //
Экологическая ординация сообществ. -М.: Наука, 1990.
-С. 79-112.
Птицы России и
сопредельных регионов. -М.: Наука, 1993. -365 с.
Птицы Советского
Союза. -М.: Сов. наука, 1951 – 1954. -Т. I – VI.
Птицы СССР. -М.; Л.: Наука, 1982, 1987 – 1989. Т. I – IV.
Пузаченко Ю.Г. Географическая изменчивость обилия и структуры
населения птиц лесных биоценозов // Орнитология. –М.:
Изд-во МГУ, 1967. –Вып.8. -С.109-122.
Равкин Е.С., Челинцев Н.Г.
Методические рекомендации по комплексному маршрутному учету птиц. – М.., 1990.
-33 с.
Равкин Ю.С. К методике учета птиц лесных ландшафтов // Природа
очагов клещевого энцефалита на Алтае. –Новосибирск:
Наука. Сиб. отд-ние, 1967. -С. 66-75.
Равкин Ю.С. Птицы
Северо-Восточного Алтая. -Новосибирск: Наука. Сиб.
отд-ние, 1973. -375 с.
Равкин Ю.С.
Пространственная организация населения птиц лесной зоны (Западная и Средняя
Сибирь). -Новосибирск: Наука, Сиб. отд-ние, 1984. -264
с.
Равкин Ю.С., Лукьянова И.В. География позвоночных южной тайги
Западной Сибири. –Новосибирск: Наука. Сиб. отд-ние,
1976. –360 с.
Сметанин В.Н. Птицы
Южного Забайкалья (пути формирования фауны и пространственная структура населения):
Авторефе. дис. ... канд. биол. наук. -Новосибирск,
1989. -22 с.
Степанян Л.С. Конспект
орнитологической фауны СССР. -М.: Наука, 1990. -727 с.
Торопов К.В., Фомин Б.Н., Козлов
Н.А. и др. Лесостепь Обь-Иртышского междуречья и долины Оби //
Пространственно-временная динамика животного населения (птицы и мелкие
млекопитающие). -Новосибирск: Наука. Сиб. отд-ние,
1985. -С.67-79.
Трофимов В.А., Равкин Ю.С.
Экспресс-метод оценки связи пространственной неоднородности животного населения
и факторов среды // Количественные методы в экологии животных: Тез. докл. -Л.: Наука. Ленингр. отд-ние, 1980. -С. 135-138.
Фертикова Е.П.
Географическая изменчивость параметров годового цикла сезонных явлений серой
славки (Sylvia communis Latham): Автореф. дис. … канд. биол. наук. –СПб.,
2000. -18 с.
Харвей Д. Научное
объяснение в географии. М: Прогресс, 1974- -502 с.
Цыбулин С.М. Птицы
диффузного города. -Новосибирск: Наука. Сиб. отд-ние,
1985. -169 с.
Шмидт-Ниельсен К. Размеры
животных: почему они так важны? -М.: Мир, 1987. -259
с.
Юдкин В.А.
Территориальная изменчивость летней динамики обилия птиц // Миграции птиц в
Азии. -Новосибирск: Наука. Сиб. отд-ние, 1986а. -С. 252-260.
Юдкин В.А. Птицы
таежного Прииртышья (пространственная организация летнего населения): Дис. ...
канд. биол. наук. -Новосибирск, 1986б. -288 с.
Юдкин
В.А., Вартапетов Л.Г., Козин В.Г., и
др. Материалы к распространению птиц
в Западной Сибири // Материалы к распространению птиц на Урале, в Приуралье и
Западной Сибири. -Екатеринбург, 1997. -С. 172-181.
Юрлов А.К., Михантьев А.И.,
Селиванова М.А. Влияние
факторов среды на продуктивность популяций околоводных птиц на юге Западной
Сибири // Сиб. Экол. Журн. –1994. –№4. -С.347-353.
Baker
M.C., Mewaldt R.L. The use of
space by White crowned sparrows: juvenile and adult ranging patterns and home
range versus body size comparisons in an avian granivore community //Behav. Ecol. a Sociobiol. 1979. -Vol.6, №1. -P. 45-52.
Blackburn
T.M., Harvey P.H., Pagel M.D. Species
number, population density and body size relationships in natural communities
// J.Anim.Ecol. -1990. –Vol.59, №1. -P.335-345.
Bruderer
B. The course of bird migration
in
Harrestad
A.S., Bunnell F.D. Home range
and body weight – a re-evaluation // Ecology. -1979. -Vol.60, №2. -P. 389-402.
Kendeigh
S.C., Dolnik V.R., Gavrilov V.M. Avian energetic // Granivourus birds in ecosystems/Eds.
G. Pinowski, S.C. Kendeigh: Intern. Biol. Programs,
Kurlavicius P. Birds of forest islands in South-East Baltic region. -
Lasiewski R.C., Dawson W.R. A re-examination of the relation
between standard metabolic rate and body weight in birds.//Condor. 1967.
-Vol.69, №1. -P.13-23.
MacArthur
R., Recher H., Cody M. On the
relation between habitat selection and species diversity // Amer. Natur. -1966,
-Vol.100, №913, -P. 319-325.
McClure
H.E. Migration and survival of the
birds of
Monjeau
A., Birrey E., Phillips C., Sikes R. Small mammals response to the hierarchical nature of landscape structure
// Euro-American Mammal Congr., Santiago de Compastela, Spain, July 19-24,
1998. -P. 23.
Ricklefs
R.E. Geographical variation in
clutch size among passerine birds: Asmole's hypothesis // Auk. 1980. -Vol.97.
-P. 38 43.
Robbins
C.S., Sauer J.R., Greenberg R.S., Droege S. Population declines in North American birds that migrate to the
neotropics // Proc. Nat. Acad. Sci. USA. -1989, -Vol.86, №19. -P. 7658-7662.
Schoener
T.W. Sizes of feeding territories
among birds // Ecology. -1968. -Vol 49. -P. 123-141.