УДК: 591:9
ОПЫТ КАРТОГРАФИРОВАНИЯ НАСЕЛЕНИЯ ЖИВОТНЫХ
Ю.С.Равкин,
Е.С.Равкин
Институт систематики и экологии животных
СО РАН
630091,
Новосибирск, ул.Фрунзе 11
МосНПО «Радон», 127562, Москва, Алтуфьевское шоссе, 30-29
Поступила
в редакцию
Описаны
приёмы, использованные при составлении карт животного населения, легенды
которых выполнены по классификациям сообществ с помощью методов кластерного
анализа по коэффициентам сходства. Описана специфика таких карт и отличия их от
составленных на экспертной основе.
ВВЕДЕНИЕ
Карта
считается началом и концом любого географического исследования [4]. В
зоогеографии ландшафтная или геоботаническая карты используются сначала в
качестве основы для сбора материала, а в завершении, карта животного населения
отображает итоговые концепции, полученные в процессе работ. Таким образом,
карта животного населения представляет собой окончательный, хотя и не
единственный результат зоогеографического исследования, которое может
ограничиваться классфикационными построениями или отображаться в виде графов и
ординационных схем в многомерном факторном пространстве. Нередко они информативнее
карт, лучше иллюстрируют причины неоднородности сообществ или её корреляции с
факторами среды. Но графы и классификации, включающие факторное объяснение
неоднородности сообществ не несут сведений о пространственной смежности
различных таксонов классификации и о площади, занимаемой ими. Картографическое
отображение территориальной изменчивости населения, особенно на больших
площадях, отражает эти особенности и является наиболее наглядной формой их
представления.
Результаты
типологической классификации животного населения нередко существенно расходятся
с ландшафтными и геоботаническими построениями. Так, на выполненной нами карте
орнитокомплексов Западно-Сибирской равнины, в соответствии с классификацией,
чётко видны две широтные полосы к северу и югу от границы между северной и
средней тайгой [20]. На карте растительности такие полосы соответствуют
подзональным отличиям и их значительно больше, чем на карте сообществ птиц.
Правда, последние отличия хорошо видны на графе и лишь при совместном анализе
орнитокомплексов Запад-но-Сибирской и Восточно-Европейской равнин отображение
на карте стало совершенно необходимым. Это связано с тем, что граница северной
и южной (срединной) надтиповых группировок в европейской части смещается к северу
фактически ещё на одну подзону из-за отепляющего влияния Северо-Атлантического
течения. Поэтому описание графа становится громоздким и недостаточно наглядным,
в то время как на карте это смещение прослеживается без особого труда.
Уникальность
территорий и распределения видов животных создаёт ряд проблем при
зоогеографическом картографировании. Индивидуальный и топологический подходы,
отражающие эту уникальность, осуществимы лишь по отдельным видам, на
ограниченных площадях и только в крупном масштабе. Во всех остальных случаях
целесообразно использовать обобщённые, в том числе типологические представления
о среде и отражать на картах неоднородность сообществ некоторых систематических
групп животных или таксонов других классификаций (экологических, хозяйственных
и др.). Обычно в такие таксоцены входят виды с существенно различающимися
экологическими требованиями к среде, что порождает необходимость отделения
главных отличий сообществ от второстепенных, поскольку отразить их полный набор
невозможно.
Для этого
выявляются виды, характерные для определённых территорий. Сначала
классифицируют виды по сходству распределения, выявляя при этом предпочитаемые
ими природно-географические подзоны и/или азональные ландшафты [18, 37].
Последние и отражаются на карте с перечнем в легенде преобладающих характерных
и сопутствующих видов с учётом их трофики и динамики численности. Этот подход
оправдан при составлении мелкомасштабных обзорных карт по населению, представленному
небольшим числом видов. Однако в этом случае возможны ситуации, когда на
отдельных территориях на долю характерных видов приходится меньшая часть
населения и эти варианты более сходны с сообществами местообитаний, где
доминируют иные характерные виды. Поэтому тип преференции, например
лугово-болотный, может быть, но на территории лугов и болот преобладают виды,
характерные для лесов [23, 24]. Поэтому распределение типов населения и видов
может не совпадать. Априорная заданность предпочтения зон, подзон или азональных
биоценозов может существенно исказить границы в неоднородности населения, чётче
выявляющиеся при более мелком ранге рассмотрения. Второй подход к сокращению
отражаемого разнообразия – это классификация животного населения не по
отдельным видам, а по эмерджентным свойствам сообществ, т.е. свойствам
комплекса в целом, не сводимого к простой сумме его частей. Эти свойства
отражаются парными коэффициентами сходства.
Оба подхода
отражают глубинные различия в понимании того, что такое население. Большинство
зоогеографов не сомневается в том, что население это территориальное сообщество
животных, связанных между собой непосредственно или опосредованно через другие
виды и/или совместным использованием территории и ресурсов, размещённых на ней.
Исследования сообществ в целом, т.е. всех животных, практически не реализуемы,
поэтому всегда изучаются лишь части его. При этом системный подход к изучению
целого и частей возможен до тех пор, пока в той или иной мере сохраняется их
целостность. Естественно, что степень эмерджентности может существенно
различаться в системах и подсистемах согласно их ступенчатой иерархии и
последовательной встроенности друг в друга.
Тем не
менее, одни исследователи тяготеют к видам и могут обойтись без явного использования
общих характеристик сообществ, другие оперируют комплексами (наборами,
статистическими ансамблями), проводя анализ пространственных изменений
таксоценов по их интегральному сходству и различиям. В первом случае важнее
всего распределение видов, в другом – пространственная дифференциация
сообществ, изменчивость облика населения в целом. Облик сообщества - это
количество и соотношение особей всех видов, зарегистрированных в выборке по анализируемым
подразделениям территории. В итоге при первом подходе виды в населении обычно
признаются равноценными и равнозначными признаками. Для этого все показатели
могут соотноситься с максимальным, средним или суммарным обилием по каждому
виду в отдельности [19]. Близкий результат может достигаться шкалированием
эмпирических показателей обилия каждого вида по отношению к нормальному, с
точки зрения исследователя, уровню его численности [13]. В итоге население
характеризуется соотношением этих долей или балльных оценок преференции вида
или по шкалам относительно нормального для данного вида уровня обилия или
биомассы. Во втором случае используется соотношение в плотности населения или
других суммарных показателей в данном местообитании. Дальнейшее развитие
первого из этих направлений в соответствии с системой ценностей приводит к всё
большему дроблению групп видов так, чтобы отразить особенности распредления как
можно большего их числа [16, 33, 34]. Во втором направлении, наоборот, проявляется
стремление к одновременному картографированию населения максимального числа
видов животных [2].
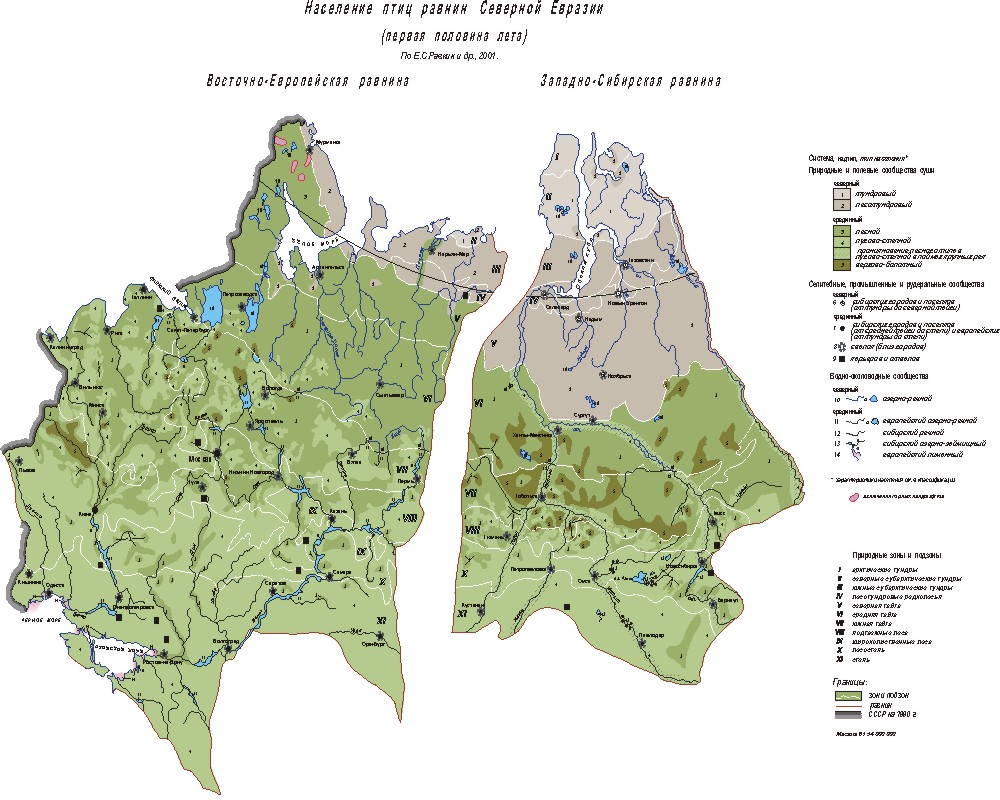
Население птиц Восточно-Европейской (а) и
Западно-Сибирской (б) равнин. Система природных и полевых сообществ птиц суши.
Северный надтип населения. Типы населения: 1 – тундровый; 2 – лесотундровый.
Срединный надтип. Типы: 3 – лесной; 4 – лугово-степной; 4а – проникновение
лесного типа в лугово-степной в поймах крупных рек; 5 – верхово-болотный.
Система селитебных, промышленных и рудеральных сообществ птиц. Северный надтип
населения. Тип населения: 6 – сибирских городов и посёлков от тундры до северной
тайги включительно. Срединный надтип. Типы: 7 – сибирских городов и посёлков
(от средней тайги до степи) и всех европейских; 8 – свалок (близ городов); 9 –
карьеров и отвалов. Система вводно-околоводных сообществ птиц. Северный надтип
населения. Типы населения: 10 – озёрно-речной тип. Срединный надтип. 11 –
европейский озёрно-речной; 12 – сибирский речной; 13 – сибирский
озёрно-займищный; 14 – европейский лиманный; 15 – включения горных ландшафтов.
Границы: 16 – зон и подзон; 17 – равнин; 18 – СССР на 1990 г. Природные зоны и
подзоны: I – арктические тундры; II – северные
субарктические тундры; III – южные субарктические
тундры; IV – лесотундровые редколесья; V –
северная тайга; VI – средняя тайга; VII – южная тайга; VIII
– подтаёжные леса; IX – широколиственные леса; X –
лесостепь; XI – степь. Характеристики населения и цветной
вариант карты опубликованы ранее. * – в оригинале отличаются цветом.
КЛАССИФИКАЦИЯ
НАСЕЛЕНИЯ КАК ОСНОВА ДЛЯ СОСТАВЛЕНИЯ ЛЕГЕНДЫ КАРТЫ
Каждая
карта – это, прежде всего, отражение какой-либо концепции о неоднородности
картографируемого объекта [5]. Поэтому разработка концепции – один из основных
этапов картографирования. Классификация как результат, по сути, и есть итоговая
концепция, правда только в том случае, когда деление на таксоны так или иначе
объяснено, причинно или коррелятивно, например, прослежена связь с некоторым
набором внешних и внутренних структурообразующих факторов. Классификация сообществ
как метод группировки вариантов населения по тем или иным признакам разработана
сравнительно хорошо. В простейших случаях процесс классификации сводится к
упорядочению по убыванию или возрастанию показателя, принятого в качестве
признака классификации: плотности населения, видового богатства, разнообразия и
т.д. Далее проводится поиск естественных разрывов в этом ряду значений. При
отсутствии таковых границы в ряду выделяются условно. Если классификация
проводится последовательно по ряду признаков, тогда менее значимый из них
служит основанием для доразбивки выше стоящего таксона на более мелкие.
Проблема, в данном случае, возникает в обосновании иерархии значимости
признаков. Поскольку доказать это в вероятностных системах очень трудно,
нередко принятая иерархия никак не обсуждается и не обосновывается и потому произвольна
(субъективна).
Возможно
построение легенд и карт исходно концептуальных, т.е. построенных исходя из
неких представлений, сформировавшихся у автора в процессе логических
размышлений по поводу информации, почерпнутой из литературы, устных сообщений
коллег и обсуждений с ними возникающих проблем, а также собственного опыта и
имеющихся сведений. Так, например, составлена карта населения птиц и
млекопитающих лесной и лесостепной зон Сибири [29]. В атласе «Алтайский край»
первое деление на орнитогеографической карте совпадает с отличиями с одной
стороны группировок населения птиц равнинных и Салаирского кряжа, с другой –
Горного Алтая [9]. Если не подозревать, что это деление проведено по группам
исполнителей проводивших работы, то логичнее деление должно быть иным: на
равнинные группировки и горные, хотя последние могут делиться на две подгруппы
– алтайских и салаирских вариантов.
Анализ
населения птиц лесной зоны Западной и Средней Сибири и Северо-Восточного Алтая
по сходству показал, что, прежде всего, имеется три системы населения птиц –
незастроенной суши, застроенной суши и водно-околоводных сообществ [22]. Вторая
система на рассматриваемой карте атласа «Алтайский край» [9] не представлена
из-за отсутствия данных, а третья включена в качестве населения основного
местообитания, объединяющего долинные лесные и водно-болотные сообщества.
Последнее, видимо, сделано для единообразия с ландшафтным делением территории,
в то время как анализ по сходству показывает, что лесные долинные (даже
пойменные) варианты населения, как горные, так и равнинные, ближе к сообществам
плакорных лесов. Также сходны между собой сообщества лугов и болот пойменных,
надпойменных и междуречных. Население водно-околоводных местообитаний и суши
резко отличается даже в пределах надпойменных террас долин крупных рек и их
пойм.
Все наши
построения получены только по имеющейся выборке данных и формализованного их
анализа. Кроме исходных концептуальных позиций, используемых при сборе и
обработке материала, карта строится по итоговой концепции, сформулированной в
результате анализа. Так, при исследовании летнего населения птиц северных
предалтайских равнин, Северо-Восточного, Северного, Центрального и
Юго-Восточного Алтая по сходству орнитокомплексов в среднем по экосистемам
(согласно авторского макета карты В.П. Седельникова масштаба 1:500000)
прослеживается деление на семь типов сообществ [27]. В первый вошло население
скал, курумов и осыпей высокогорья; во второй – высокогорных тундр и
тундростепей; в третий – настоящих и опустыненных степей высокогорий и
среднегорий аридных провинций; в четвёртый – лесов, редколесий, лесостепных,
пойменных и болотных комплексов; в пятый – лугов, полей и степей предгорно-низкогорных
и среднегорных ландшафтов гумидных провинций; в шестой – селитебных территорий;
в седьмой – рек и озёр.
Деление
населения мелких млекопитающих в атласе «Алтайский край» аналогично
орнитогеографическому [11]. Сравнивать его труднее, т.к. наши материалы собраны
канавками и поэтому включают не только грызунов, но и землероек, а информация
по зайцеобразным у нас отсутствует. Самое общее (первое) формализованное
деление по этим данным позволяет говорить о шести типах населения. В первый из
них входят сообщества тундр, степей и тундростепей аридных высокогорий; во
второй – тундр гумидных высокогорий; в третий – редколесий, темнохвойных и
темнохвойно-лиственных лесов, посёлков, а также лесостепных и пойменных
комплексов; в четвёртый – светлохвойных, мелколиственно-светлохвойных и
мелколиственных лесов; в пятый – предгорных и низкогорных лугов, полей и
луговых степей; в шестой – настоящих и опустыненных степей.
Таким
образом, формализованное деление животного населения по коэффициентам сходства
может существенно отличаться от экспертно-умозрительного.
При составлении
карты населения птиц СССР, Нижнего Приамурья и Камчатской области [6–8, 10]
проведено разделение орнитокомплексов на уровне типа по преобладанию видов
одной крупной географо-генетической или экологической группы
(южнотаёжно-нетаёжной, северотаёжно-среднетаёжной или лесной, кустарниковой и
т.п.) и/или по сходству видового состава в группах с разными доминантами. При
отсутствии явного доминирования видов той или иной группы, выделялись переходные
варианты. Недостатками такого подхода следует считать произвольность
(необоснованность) иерархии признаков классификации; неоднозначность подходов к
выделению типов населения; использование доли вместо обилия (сходные доли могут
иметь виды с существенно различной численностью); оценку сходства по
принадлежности к группе. Использование этих приёмов, так же, как переход на
обобщённые характеристики (знаки различного «веса»), целесообразно, когда
оценки обилия животных ненадёжны из-за недостаточной протяжённости маршрутов,
имеют существенные методические или индивидуальные отличия при сборе, а также
при существенных годовых колебаниях обилия.
Процедура
«расстригания» таблиц на горизонтальные и вертикальные полосы по сходству в
расположении и весу знаков, символизирующих долю в населении [8], применима
лишь к небольшим выборкам, в то время, как использование для классификации
вычислительной техники, в этом плане практически не имеет ограничений,
существенно облегчает работу и делает результаты менее субъективными. В
принципе, оба приёма должны давать сходные классификации, если они выполнены по
доле, а не по обилию.
Использование
коэффициента сходства для количественных признаков, в качестве единственного
критерия для классификации, делает необязательным в легенде отражение
представленности различных экологических и фауно-генетических групп, в том
случае, когда отражаемое ими явление значимо прослеживается в населении в виде
зонально-подзональных или провинциальных таксонов классификации [20, 25]. Это
упрощает легенду и возможность её восприятия, что существенно при составлении
карт, выполненных в относительно крупном масштабе на значительные территории.
Изменение доли экологических и географо-генетических групп может быть сюжетом отдельных
карт [36], или быть дополнительной характеристикой таксонов классификации
населения, но не признаком для их выделения.
Следует
отметить также, что все классификации составляются при недостатке информации,
т.е. при бóльших или меньших ошибках репрезентативности, которые
суммируются при вторичных классификациях выполненных с использованием первичных
умозрительных обобщений. Кроме того, всякая классификация есть получение нового
знания за счёт обобщения и одновременно, по причине огрубления представлений, потеря
индивидуальной информации, в данном случае о распределении видов. Такое
огрубление может породить ошибочные суждения. Так, в упомянутой
географо-генетической классификации В.В. Брунова имеется группа
северо-среднетаёжных видов. Объединение их приводит во вторичной классификации
к затушёвыванию границы между сообществами северной и средней тайги [25]. В
результате среднетаёжное население птиц на карте В.В.Брунова [7] оказалось
ближе к северотаёжному, а не к сообществам южных подзон, как в наших
классификациях по Западной Сибири. Причины этих различий, видимо, в групповом и
видовом уровне оценки сходства и, возможно, в недостаточности выборки к моменту
классификации видов В.В.Бруновым.
Поэтому при
картографировании населения лучше всего использовать наиболее дробный уровень
рассмотрения (видовой) и единственный классификационный признак, отражающий
следовое влияние всех факторов среды (в прошлом и настоящем), а также
последствия внутринаселенческих взаимоотношений и процессов. Таким признаком
может служить сходство-различие вариантов населения по любому показателю
(обилию, биомассе, энергетике каждого вида).
Оценкой
неоднородности облика сообществ выступает матрица парных коэффициентов сходства
вариантов населения. Эта парность сравнений каждого варианта относительно
других сообществ и делает матрицу многомерной. Каждое значение в ней несёт
информацию о данном сообществе относительно другого, а строки (столбцы) в
матрице в целом, отражают сходство каждого варианта со всеми остальными. Формально
любая классификация строится по сходству в признаках, принятых в качестве
основания для деления. При использовании коэффициентов сходства, за счёт
жёсткого закрепления его меры и процедуры сравнения объединение в группы менее
субъективно, так как выбор исследователя относится только к мере общности и способу
классификации, но полностью снимает субъективизм в иерархии значимости признаков.
Классификации
животного населения, выполненные с одной стороны по коэффициентам сходства, а с
другой, сначала по общности доминантов, а потом по доминированию типов фауны,
обычно дают близкие результаты. Это обусловлено тем, что в обоих случаях значимое
сходство определяется преобладающими видами. Но в первом случае оно определяется
по абсолютным или относительным значениям обилия [21], а во втором по доле в
населении. Поэтому возможны случаи, когда при оценке сходства по доле
доминантов в один и тот же тип войдут варианты с одинаковым составом лидеров,
но резко различающиеся по уровню обилия, как суммарного, так и преобладающих
видов. По коэффициентам общности сходство этих вариантов будет незначительным.
Полученное
так или иначе разбиение необходимо объяснить факторами среды или особенностями
внутринаселенческих отношений. Эта процедура фактически сводится к поиску корреляций
неоднородности сообществ и среды. Предварительная автоматическая классификация
по сходству упрощает поиск этих факторов. В дальнейшем они служат основанием
для экстраполяции на необследованные выделы карты местообитаний, а также
адресом для выявления границ размещения таксона на местности.
Этот
подход, в отличие от принципа картографирования населения вместе с его
местообитанием [35] носит сугубо населенческий характер, так как условия среды
служат объяснением классификации, но не её признаком. Однако результат такого
деления многим кажется простым разнесением характеристик населения по таксонам
геоботанической или ландшафтной классификации. Возможно, это связано с тем, что
названия населенческих таксонов даются обычно по коррелирующим с растительностью
признакам (например, лесной, лесополевой или низинно-болотный тип сообществ).
Однако лесной тип, например, отнюдь не совпадает с распространением только
лесов и тем более всех из них. Так, на Западно-Сибирской равнине в него входят
сообщества птиц лесов, облесённых низинных и переходных болот, зарастающих
вырубок, гарей и полей, чередующихся с перелесками, если доля последних по площади
больше, чем полей. Распространение этого типа ограничено пространством от
степной зоны до средней тайги включительно, а аналогичные северотаёжные
варианты, так же как и лесотундры, имеют лесотундровое население. Таким
образом, название "лесной" отражает дендрофильную специфику
орнитокомплексов в пределах насаждений нормальной полноты, но не на всей
занятой ими территории.
Нам кажется
менее удачным названия типов животного населения по характерным видам или их
группам. Дело не только в их непривычности. Названия по признакам среды,
богаче, т.к. одновременно дают адресную и экологическую (адаптационную)
характеристику населения, избавляя от необходимости давать её в качестве
дополнительной информации. Название типа в виде перечня преобладающих видов с указанием
распределения [32], воспринимается легче, но оно не компактно. Так, название
одного из типов в указанной публикации звучит как: «Степные пеструшки и суслики
в зональных и горных степях». Этот тип можно назвать и пеструшково-сусликовым и
степным. Последний вариант как маркёр проще воспринимается и привычнее, а
доминанты (лидеры) и перечень местообитаний даются как дополнительная
характеристика. В первом и втором варианте видовая специфика населения
представлена на первом месте, а территориально-экологическая на втором или
последнем. В значительной степени это дело вкуса и привычки, хотя, в какой-то мере,
это и проблема восприятия и воспроизведения названий, т.е. имеет и дидактическое
значение.
Л.Г.Емельянова
и А.К.Даниленко, В.Ю.Румянцев использовали сопоставление животного населения
соседних выделов в пределах трапеций карты-основы или соседних местообитаний [14,
15]. Если характеристики населения сходны, выделы и характеристики их населения
объединялись, если нет, то выяснялись причины этих отличий. Если авторы считали
отличия достоверными, то варианты населения оставляли в качестве самостоятельных
выделов. Л.Г. Емельянова, кроме того, рассчитывала по данным, собранным
ограниченное число лет, максимально возможное обилие каждого вида грызунов. В
принципе, такие подходы вполне правомерны, но едва ли обоснованы, т.к. оценка
достоверности различий проводилась экспертно, а не статистически. Последнее
требование может быть выполнено при значительных объёмах материала, которых,
как правило, в распоряжении авторов нет.
По
границам, выявленным по коэффициентам сходства на карте, использованной в
качестве основы, оконтуривается обобщённый выдел и его подразделения,
соответствующие распределению таксонов населения. Таким образом, карта
животного населения представляется результатом генерализации и экстраполяции
или деления выделов карты-основы с использованием других картографических
источников. Основанием для генерализации, экстраполяции и деления выделов
служат факторы среды, одинаково проявляющиеся в совокупности выделов со
сходным, по принятым принципам, животным населением.
Типологический
подход, как уже говорилось, даже при картографировании не отражает с должной
полнотой хорологические связи сообществ, иерархию использования животными
территории, т.к. при динамической оценке обилия одни и те же особи по времени
нахождения могут относиться к существенно разным таксонам классификации. Хорологическое
выделение минимальных территорий и объединение выделов карты местообитаний в
более крупные подразделения, которые проводятся по ширине арены
жизнедеятельности животных, отражают эту иерархию, однако отсутствие жёсткого
алгоритма такого объединения препятствует повторяемости и однозначности
решений, приближая такие работы к искусству единомышленников. Заданность границ
населения субъектом исследования переносит процедуру объяснения на период
работы после создания картографического продукта. При предварительном типологическом
анализе карта лишь отражает, иллюстрирует его результаты. Однако это не
исключает продолжение анализа по составленным картам и получения дополнительной
информации с их помощью.
СПОСОБЫ
КОМПЕНСАЦИИ РАЗЛИЧИЙ В ОБЪЁМАХ СОБРАННОГО МАТЕРИАЛА
Случаи
специального сбора данных для картографических целей очень редки. Обычно
используется весь объём имеющихся материалов как опубликованных, так и
фондовых, включая собранные в разные годы, разными исполнителями, в разном
объёме и с различиями в методиках. Использование таких материалов при
классификации нередко сопровождается резкими незакономерными отклонениями
отдельных проб от общих тенденций. При автоматической классификации эти
выпадающие варианты выделяются в виде самостоятельных групп, в то время как более
общие закономерные отличия на их фоне, фигурируют как второстепенные и
незначимые. Кроме того, на результатах нередко сказывается несбалансированность
выборки. Так, по птицам большая часть учётов обычно проводится в лесах, а в
открытых, особенно в антропогенных местообитаниях и на болотах, значительно
меньшая. Это связано с неинтересностью для орнитологов подобного населения или
с трудностями прохождения таких ландшафтов. Подобная несбалансированность при
классификации приводит к большей детализации в подвыборках, представленных
значительным числом проб.
Для
устранения этих недостатков перед классификацией можно усреднить все данные по
выделам карты-основы, например, местообитаний или растительности. Тогда выборка
будет типологически соответствовать неоднородности сообществ в ранге наименьшей
единицы рассмотрения. Впервые этот приём использован Е.М. Снигиревской [30].
Н.В.Тупикова [31] считает, что это равнозначно априорному допущению
однотипности населения внутри растительной формации. Однако это вовсе не обязательно
и может сводиться к утверждению, что внутри наименьшей единицы, они просто не
рассматриваются, так же как и годовые колебания.
Достаточно
полная для картографирования информация о населении обычно имеется лишь по
отдельным таксоценам и нередко сведения о численности приводятся в пересчёте на
разные единицы: площади, расстояния, времени или орудия лова. Последнее делает
невозможным прямые расчёты сходства для разных видов.
Есть
несколько способов компенсации таких недостатков. Первый из них сводится к
декларации равенства всех видов как признаков классификации независимо от их
обилия. Это достигается нормированием показателей по максимальному, среднему
или суммарному значению показателей. Тогда значимым оказывается только характер
распределения каждого вида, а не уровень его численности по отношению к обилию
остальных видов, отмеченных на исследуемой территории. Это отражает не
неоднородность населения, а только предпочитаемость территории видами,
естественно в пределах возможного. Построенная так классификация, легенда и
карта должны называться не населенческими, а преферентными. В противном случае
мы должны согласиться, что плотность населения для нас не значима и одинаковыми
следует считать сообщества на территориях со сходным соотношением в степени
привлекательности их для животных, но с существенным различием в соотношении
показателей обилия слагающих его видов. Например, максимальное обилие по весьма
многочисленному и очень редкому виду будут иметь одинаковый вес, скажем 100%.
Использовать этот приём можно для доразбивки групп на подгруппы [28]. В этом
случае деление проводится по ненормированным показателям, пока это даёт
интерпретируемый результат, а затем по нормированным для дополнительного
разбиения. Характеристики населения в этом случае даются по исходным данным.
Это вовсе
не значит, что классификации и карты, составленные по нормированным или
шкалированным значениям, не нужны или их реализация порождает артефакты. Просто
они отражают относительный внутривидовой уровень в распределении видов.
Балльное шкалирование проводится по отношению к экологической норме видового
обилия [12, 13]. К сожалению, выявление этой нормы и отклонений от неё
возможно, как пишут и сами авторы, не по всем видам и весьма субъективны. Такой
подход служит, кроме прочего, одним из эффективных способов компенсации
недостатков используемых материалов и размерных отличий животных, однако он
приводит к формированию иных представлений о населении, связанных не с
формальным числом особей на единицу площади, а с оценкой различий территорий по
степени их привлекательности для животных и возможности использования, в том
числе при конкуренции.
Некоторые
сомнения вызывает и утверждение, что низкая численность всегда свидетельствует
о неустойчивости воспроизводства. В действительности нередки случаи, когда это
связано с малой площадью благоприятных участков внутри крупного, но в целом
менее благоприятного выдела. На таких вкраплениях эффективность размножения и
локальное обилие столь же велики, как и на оптимальных территориях. Эффект низкой
численности определяется усреднением по этим различным по привлекательности частям
общего выдела. К преферентным следует отнести и карту "Биоразнообразие
населения наземных позвоночных России" [13]. Её лучше называть картой
преферентно-видового богатства населения.
Сомнение
вызывает утверждение, что такие карты информативнее, чем инвентаризационные.
Такое мнение справедливо лишь для карт, не обеспеченных фактическими материалами.
Сама идея введения баллов (групп) обилия по отношению не к формальным границам
[17], а к уровню естественной (средней, суммарной, максимальной, нормальной) численности
широко использовалась в классических эколого-фаунистических исследованиях.
Всё таки
следует приводить все показатели к единой пересчётной единице, насколько это
возможно. Если это удаётся сделать (хотя бы примерно) по большей части
картографируемого таксона, то классификацию можно выполнить по этим видам,
поскольку для них имеются сопоставимые данные. Так, в Атласе Новосибирской
области [2] классификация выполнена одновременно по населению земноводных, птиц
и мелких млекопитающих, для которых имелись показатели обилия в пересчёте на 1
км2. На долю этих групп животных приходится 88-98% обилия всех позвоночных
суши. Поэтому даже если бы была сопоставимая информация по оставшимся крупным и
средним млекопитающим и пресмыкающимся, классификационное разбиение населения
не изменилось бы. В этом случае, характеристики таксонов классификации
населения приводятся дифференцированно по обеспеченной и не обеспеченной части
сообществ. Так, сведений по обилию промысловых млекопитающих в ландшафтных
урочищах не было, но имеющейся информации достаточно для характеристики в
рамках типа и подтипа населения. В принципе, также можно использовать данные,
приведённые к разным единицам пересчёта, т.е. по обеспеченным материалами видам
в легенде приводятся средние значения в пересчёте на единицу площади, а по
остальным – относительные показатели, в пересчёте на береговую линию, ловушки и
т.п.
Тот же
подход использован при составлении карты по численности животных, включённых в
Красную книгу Новосибирской области [2]. Обычно же при картографировании
исследователи предпочитают равняться на нижний уровень информационной
обеспеченности или ограничивать таксоцен только теми видами, по которым имеется
достаточная информация. По предлагаемому способу на карте отражается вся имеющаяся
информация.
КОМПЕНСАЦИЯ
РАЗЛИЧИЙ В РАЗМЕРНОСТИ И ЧИСЛЕННОСТИ ЖИВОТНЫХ
Известно,
что численность видов, мелких по размерам, обычно выше, чем крупных животных.
Поэтому, в случае классификации населения по обилию, выявленные тренды будут
свойственны, в основном, мелким животным, в то время как роль одной особи в
биоценозах зависит от её массы и, соответственно, от количества
трансформируемой ею энергии. Так, в населении наземных позвоночных
Новосибирской области преобладают земноводные. Однако интенсивность потока
энергии, проходящего через их популяции, значительно меньше, чем более крупных
теплокровных животных. Кроме того, распределение сеголеток земноводных, доля
которых во второй половине лета очень велика, существенно зависит от близости
канавок к водоёмам с успешным выплодом [26]. Информации об этих водоёмах на
снимках и картах нет, поэтому распределение сеголеток вносит элемент
неопределённости в распределение земноводных и в целом позвоночных суши. Это
осложняет интерпретацию изменений сообществ в пространстве.
Для
компенсации такой диспропорции в собранных материалах, можно перед
классификацией перейти на энергетические показатели. Тогда полученное разбиение
на группы будет больше соответствовать биоценотической значимости животных и
зависимости их от неоднородности среды, чем формальное число особей. В
характеристиках населенческих таксонов, по крайней мере, позвоночных, после
классификации в легенде карты можно вернуться к значениям обилия, как к более
привычным и понятным. То есть классификация проводится по энергетическим
показателям, а выделенные таксоны характеризуются числом особей на единицу
площади. Однако это не всегда приводит к сбалансированному отражению
неоднородности сообществ по всем классам позвоночных, хотя желание иметь его в
легенде весьма велико. Выходом из этой ситуации могут быть характеристики,
приводимые отдельно по классам позвоночных и любым выделенным группам, например
животным, включённым в Красные книги [7] или охотничьим и прочим видам, по
которым есть информация лишь о присутствии-отсутствии. Информативность такой
легенды гораздо выше, хотя все сведения даются в рамках единой комплексной
классификации.
Особую
несовместимость могут иметь данные по рыбам и наземным позвоночным. Так, по
Новосибирской области для рек и водоёмов имеется материал лишь по
рыбопродуктивности, т.е. суммарной массе рыб в уловах. Чёткая пространственная
разобщённость водных экосистем и суши позволяет отразить их население на одной
карте в виде несвязанных самостоятельных классификаций, отражённых в легенде.
Это приводит к экономии места и более подробной характеристике населения описываемой
территории на одной карте. По рыбам классификация выполнена экспертно по
продуктивности и преобладающим видам, а по наземным позвоночным
формализованными методами. Фактически это совмещение в одном изображении двух
разных карт.
По
Новосибирской области удалось совместить в одной карте информацию о летнем и
зимнем населении птиц [3]. Это стало возможным благодаря тому, что границы
зимнего и летнего населения в ранге типа почти совпадали, а подтипы выделялись
только в летний период.
ОСОБЕННОСТИ
ХАРАКТЕРИСТИК НАСЕЛЕНИЯ НА КАРТАХ, ВЫПОЛНЕННЫХ ПО КОЭФФИЦИЕНТАМ СХОДСТВА
В легендах
карт, построенных по одному из суммарных показателей населения, обычно
выделяются неперекрывающиеся для каждого выдела пределы значений плотности,
видового богатства и т.д. [1]. На картах, легенды которых сформированы по
коэффициентам сходства, приводятся средние значения этих параметров, а не
диапазоны. При достаточно большой выборке можно привести и пределы
вариабельности значений, однако они всё равно будут пересекаться или будут
близкими по значимости в разных выделах. Дело в том, что в разных по облику
группах населения может быть сходная плотность, биомасса или видовое богатство.
Поскольку не они приняты в качестве дифференцирующих, их невозможно отразить в
виде неперекрывающихся диапазонов.
Характеристики
населения в легенде и, соответственно, по таксонам классификации рассчитаны как
средние по всем вошедшим в них вариантам. Поэтому и лидирующие виды относятся к
территории в среднем, а не к каждому индивидуальному выделу, при этом на их
территории часть средних лидеров может не встречаться вообще. Например, белошапочная
овсянка в среднем является первым из лидеров верхово-болотного типа населения,
распространённого как в Западной Сибири, так и в Европе, хотя этот вид на
территории последней не встречается [25].
Дробность
деления при автоматической классификации в разных таксонах может быть
различной, т.к. используются только те разбиения, которые удаётся объяснить,
т.е. связать с неоднородностью каких-либо факторов среды.
ЗАКЛЮЧЕНИЕ
В
дополнение к ранее используемых принципам зоогеографического картографирования
нами реализованы следующие приёмы. Во-первых, это совместное картографирование
населения всех позвоночных на количественной основе на одной карте. Во-вторых,
использование энергетического подхода для снятия влияния размерности животных и
географически незакономерных всплесков численности отдельных видов. В результате
подразделение животного населения в целом проводится на более надежных основаниях.
Классификация
и факторный анализ проведены по группам животных, наиболее обеспеченным
исходными данными (например, совместно по земноводным, птицам и мелким млекопитающим,
на долю которых приходится подавляющая часть населения наземных позвоночных).
По остальным группам, с меньшим объемом имеющихся материалов, сведения разносятся
по предварительно выделенным таксонам классификации сообществ. Это позволяет
при составлении легенды использовать для принятия решений о границах наиболее
надежную часть информации.
Легенда
разрабатывается на основе предварительной классификации сообществ по сходству,
с последующим поиском связей их неоднородности с факторами среды, в первую
очередь с характером растительности и другими пейзажными признаками,
оцениваемыми экспертно. Выявленные связи использованы при объединении выделов
карты-основы, имеющих сходное население в выделы карты сообществ.
При
совместном картографировании населения животных существенно отличающихся по
численности и площади арены жизнедеятельности, в легенде можно сохранить
специфику каждой из групп животных. Для этого на одной карте следует давать
информацию дифференцированно, например, по рыбам, земноводным, пресмыкающимся,
птицам и млекопитающим. Дополнительно могут быть выделены и другие группы животных
(например, по их использованию), что существенно повышает информативность
легенды. Характеристики на карте, в этом случае, отражают привычные показатели,
такие, как рыбопродуктивность или, по наземным позвоночным – плотность и
видовое богатство населения, хотя пространственное разграничение сообществ
выполнено по энергии существования животных, которая лучше отражает
биоценотическое значение видов и зависимость их от среды. Поэтому формирование
таксонов такой классификации сообществ естественнее, чем по обилию. Кроме того,
в легенде могут быть приведены лидирующие виды и другая вспомогательная информация.
При
разработке легенд предусмотрена возможность разной полноты информации по разным
группам животных и выделам карты. В тех случаях, когда отсутствуют сведения о
численности животных некоторых классов, приведены только названия лидирующих
видов, которые определяются экспертно. Это позволяет равняться не на
минимальную обеспеченность во имя единообразия легенды, а, наоборот, на самый
верхний из возможных уровень знаний дифференцированно по группам.
БЛАГОДАРНОСТИ
Авторы
искренне признательны И.Н.Богомоловой, Л.Г. Вартапетову, А.К.Даниленко,
Л.Г.Емельяновой, Т.В.Котовой, С.Г.Ливанову, Н.В. Тупиковой, Л.А.Хляп и
С.М.Цыбулину за ряд ценных советов по улучшению текста, высказанные при
обсуждении статьи. Исследования, послужившие её основой, поддержаны СО РАН в
рамках интеграционного проекта № 145.
СПИСОК
ЛИТЕРАТУРЫ
1. Атлас
биологического разнообразия лесов Европейской России и сопредельных территорий.
М., 1996, 144 с.
2.
Атлас Новосибирской области. Федеральная служба геодезии и картографии России.
М., 2002, с. 24-27.
3. Атлас юного
туриста-краеведа Новосибирской области. Федеральная служба геодезии и
картографии России. М., 1996, с. 15.
4. Берг Л.С.
Географические зоны Советского Союза. М., 1947, 398 с.
5. Берлянт А.М.
Образ пространства: карта и информация. М., Мысль, 1986, 240 с.
6. Брунов В.В.
Опыт анализа фаунистических групп птиц // Бюллетень МОИП, Отд. биол., 1978, т.
83, вып. 5, с. 5-16.
7. Брунов В.В.
Принципы и методы создания «Карты населения птиц СССР» для высшей школы //
Вестник Московского университета, сер. 5. География, 1984, № 3, с. 30-37.
8. Брунов В.В.,
Бабенко В.Г., Азаров Н.И. Население и фауна птиц Нижнего Приамурья // Птицы
осваиваемых территорий. Исследования по фауне Советского Союза (сборник трудов
Зоологического музея МГУ). М., 1988, т. 26, с. 78-109.
9. Брунов В.В.,
Даниленко А.К., Каплин В.Г., Чельцов-Бебутов А.М. Орнитогеографическая карта //
Алтайский край (атлас), 1978, М.-Барнаул, с. 128-131.
10. Брунов В.В.,
Лобков Е.Г., Баловнев В.Ю. и др. Опыт составления карты населения птиц
Камчатской области в гнездовой период // Бюллетень Московского общества
испытателей природы. Отдел биологический. 1992, т. 97, вып. 5, с. 14-26.
11. Даниленко
А.К., Кандалова Г.Т., Тупикова Н.В., Хляп Л.А., Чельцов-Бебутов А.М.
Териогеографическая карта // Алтайский край (атлас), 1978, М.-Барнаул, с.
132-135.
12. Даниленко
А.К., Куприна А.В. Принципы разработки и построения карты "Население
млекопитающих бассейна р.Алей" // Вестник Моск. ун-та. М.: МГУ, сер. V. География, 1988, № 3, с. 46-54.
13. Даниленко
А.К., Румянцев В.Ю. Биологическое разнообразие населения наземных позвоночных и
его отражение на карте // Вестник Московского университета. Серия 5, География,
1999, № 5, с. 20-24.
14. Даниленко
А.К., Румянцев В.Ю. Унификация показателей обилия наземных позвоночных России
для целей картографирования // Вестник Московского университета. Серия 5, География.
2001, № 2, с. –20.
15. Емельянова
Л.Г. Принципы и основные этапы создания карты населения мелких млекопитающих
СССР // Общая и региональная териогеография. М.: Наука, 1988, с. 310-342.
16. Емельянова
Л.Г., Назырова Р.И. Кадастрово-справоч-ная карта распространения русской
выхухоли (Desmana moshata L.,
1758)// Материалы международного совещания «Редкие виды млекопитающих России и
сопредельных территорий». М., 1997, с.102.
17. Кузякин А.П.
Зоогеография СССР. Уч.зап. Моск.обл.пед. института. Т. СIX. М., 1962, с. 3-182.
18. Конева И.В. Разработка специального
содержания карты населения грызунов и зайцеобразных Азиатской России // Картографирование
населения грызунов и зайцеобразных Азиатской России (материалы к карте). Иркутск,
1988, с. 4-39.
19. Пузаченко Ю.Г. Некоторые методические
вопросы составления мелкомасштабной карты населения млекопитающих равнин
Средней Азии // Вопросы зоологической картографии. М., 1963, с. 126-128.
20. Равкин Е.С.,
Равкин Ю.С., Вартапетов Л.Г. и др. Классификация летнего населения птиц равнин
Северной Евразии // Сибирский экологический журнал, 2001, т. 8, № 6, с.
741-766.
21. Равкин Ю.С.
Опыт картографирования зимнего населения птиц Северо-Восточного Алтая //
Вопросы зоологической картографии. М., 1963, с. 129-130.
22. Равкин Ю.С.
Пространственная организация населения птиц лесной зоны. Новосибирск: Наука,
1984, 264 с.
23. Равкин Ю.С.,
Богомолова И.Н., Ердаков Л.Н. и др. Особенности распределения мелких
млекопитающих Западно-Си-бирской равнины // Сибирский экологический журнал, №
3-4,1996, с. 307-317.
24. Равкин Ю.С.,
Богомолова И.Н., Ердаков Л.Н. и др. Пространственно-типологическая структура и
организация населения мелких млекопитающих Западно-Сибирской равнины // Успехи
современной биологии, 1997, т. 117, вып. 6, с. 738-754.
25. Равкин Ю.С.,
Вартапетов Л.Г., Юдкин В.А. и др. Пространственно-типологическая структура и
организация летнего населения птиц Западно-Сибирской равнины // Сибирский экологический
журнал, 1994, т. 1, № 4, с. 281-290.
26. Равкин Ю.С.,
Панов В.В., Вартапетов Л.Г. и др. Особенности распределения земноводных на
Западно-Сибирской равнине // Вопросы экологии и охраны позвоночных животных.
Киев-Львов, 1998, вып. 2, с. 49-77.
27. Равкин Ю.С.,
Швецов Ю.Г., Малков Н.П. и др. Плотность, биомасса и разнообразие летнего
населения наземных позвоночных Алтая // Доклады на международном симпозиуме
«Модели устойчивого социально-экономического развития Республики Алтай и стран
Алтае-Саянского региона». Горно-Алтайск, 1997, с. 20-36.
28. Равкин Ю.С.,
Сергеев М.Г., Седельников В.П. Дельта-разнообразие экосистем и информационной
стоимости биоценозов // Информационная теория стоимости и системные
экономические оценки природных ресурсов. Изд-во СО РАН. Новосибирск, 1999, с.
374-405.
29. Реймерс Н.Ф.
Птицы и млекопитающие южной тайги Средней Сибири. М.-Л.: изд-во Наука, 1966,
418 с.
30. Снигиревская
Е.М. Опыт крупномасштабного картографирования населения грызунов // Доклады АН
СССР, 1963, т. 152, № 1, 208-211.
31. Тупикова Н.В.
Картографирование животного населения // Итоги науки и техники. Серия Биогеография.
Т. 1. М., 1976, с. 98-218.
32. Тупикова Н.В.
Опыт создания карты населения грызунов (Rodentia) и пищух (Ochotona)
территории бывшего СССР: содержание и принципы построения легенды // Успехи современной
биологии. Т. 116, вып. 2, 1996, с. 243-254.
33. Тупикова Н.В.,
Комарова Л.В. Принципы и методы зоологического картографирования. М.: Изд-во
Московского университета, 1979, 191 с.
34. Тупикова
Н.В., Хляп Л.А., Варшавский В.В. Грызуны полей Северо-Восточной Палеарктики //
Зоологический журнал. Т. 79, № 4, 2000, с. 480-494.
35.
Чельцов-Бебутов А.М., Чибисова В.В. Животный мир и его ресурсы // Комплексные
региональные атласы. М.: Изд-во Московского университета, 1976, с. 326-341.
36. Штегман Б.К.
Основы орнитогеографического деления Палеарктики. Фауна СССР. Птицы. М.-Л., 1938, т. 1, вып. 2, 156 с.
37. Khlyap L.A., Warschavsky
A.A., Neronov V.M., Tupikova N.V. Biodiversity of rodents and pikas of the
Northern Eurasia (creation of GIS and analysis with using faunistic complexes)
// Biodiversity and dynamics of ecosystems in North Eurasia.