УДК 591.9(571.1)+591.526
Сезонная динамика населения птиц Среднего Урала
С.Г. ЛИВАНОВ
Институт систематики экологии животных СО РАН 630091
Новосибирск,
ул. Фрунзе, 11
АННОТАЦИЯ
Проанализированы материалы круглогодичных учетов птиц,
проведенных автором в Висимском заповеднике и его
охранной зоне с декабря 1983 по ноябрь 1984 гг. в шести местообитаниях. С
помощью классификации упорядоченных объектов выявлено 6 общих сезонных периодов:
относительной зимней стабилизации; предвесенних кочевок и начала прилета;
гнездования на фоне местных и миграционных перемещений; послегнездовых
местных кочевок и начала отлета; отлета и осеннего пролета; окончания пролета и предзимних кочевок. Приведена
краткая количественная характеристика периодов. С помощью неметрического
шкалирования выявлено 4 типа внутригодовой динамики орнитокомплексов. Сопоставление с имеющимися литературными
данными показало, что на Среднем Урале, северной лесостепи Приобья,
на Центральном Алтае и пустынях Туркмении внутригодовая устойчивость облика орнитокомплексов снижается от населенных пунктов и лесов к
полуоткрытым и открытым местообитаниям, а внутригодовая близость облика лесных орнитокомплексов снижается от южной тайги к тугаям пустынь. На Урале обилие оседлых видов сравнительно
высоко и смена аспектов населения определяется гнездящимися перелетными видами.
В лесостепи и, особенно, в тугаях доля видов,
обитающих во внегнездовое время (кочующих, пролетных,
зимующих), значительно выше, что и обусловливает усиление межсезонных отличий вариантов
населения птиц. По характеру сезонной изменчивости населения птиц, орнитокомплексы юга бореальных растительных областей и
пустынной зоны отличаются календарными сроками смены периодов. Кроме того, орнитокомплексам тугаев свойственна
большая внутригодовая дифференциация.
ВВЕДЕНИЕ
Исследования
пространственной неоднородности населения птиц в последние десятилетия получили
широкое распространение, в то время как внутригодовая динамика орнитокомплексов изучается гораздо реже [1]. Особенно мало
работ, посвященных многомерному факторному соотнесению
пространственно-временного сходства орнитокомплексов
с сезонной ритмикой природы. Это исследования, проведенные в Подмосковных смешанных
лесах[2, 3], в северной лесостепи Приобья [4], в г.
Новосибирске [5], в южной тайге Среднего Урала [6-8], в тугаях
Мургаба и Теджена зоны пустынь Туркмении [9] и на
Центральном Алтае [10]. Данное сообщение имеет своей целью отразить особенности
сезонной периодизации облика населения птиц Среднего Урала
и провести предварительные обобщения по всем имеющимся в литературе сведениям.
МАТЕРИАЛЫ И МЕТОДЫ
Методы сбора и анализа, личных и
сравниваемых в разделе «Обсуждение» опубликованных материалов, как правило,
идентичны. Расхождения, в случае необходимости, оговорены.
В работе приведены материалы круглогодичных учетов
птиц, проведенных автором в Висимском заповеднике и
его охранной зоне с декабря 1983 по ноябрь 1984 гг. (Пригородный район и
территория, приданная г. Кировграду Свердловской области).
Всего за время работ пройдено 1080 км маршрутных учетов птиц в 6 местообитаниях.
Птицы учтены на постоянных, но не строго фиксированных
маршрутах, независимо от расстояния до них от линии хода учетчика, с последующим
раздельным пересчетом на площадь по среднегрупповым
дальностям обнаружения. Для расчета обилия летящих особей вносилась поправка на
скорость перемещения [11]. За каждые полмесяца наблюдений, в каждом из
выделенных местообитаний с середины апреля по середину октября пройдено по 5 км
учетов. В остальные сезоны 1983-84 гг. протяженность маршрута увеличена в два
раза и составляла не менее 10 км для каждого местообитания за каждые полмесяца.
Таким образом, для
расчетов и сопоставления использованы 144 варианта населения птиц, характеризующие
изменчивость облика орнитокомплексов в течение года в
пихтово-еловых, пихтово-еловых с примесью березы и осиново-березовых лесах, на
свежих вырубках, сенокосах, чередующихся с перелесками, и в малых поселках.
Выяснение границ сезонных аспектов
проведено с использованием классификации упорядоченных объектов [12]. За меру
сходства облика орнитокомплексов принят коэффициент Жаккара в модификации Р.Л. Наумова [13]. Сначала для
каждого местообитания в отдельности рассчитаны хронологически упорядоченные матрицы
коэффициентов сходства 24 вариантов орнитокомплексов,
населяющих этот выдел в течение года. Затем ряд коэффициентов без права их
перестановки последовательно делился на 2, 3 и т.д. классов таким образом,
чтобы общность внутри классов была наибольшей, а сходство между ними
наименьшим. По этой программе оценивается не только сходство между пробами
соседних хронологических отрезков, но и учитывается сходство каждой пробы со
всеми остальными. Это снижает вероятность проведения ошибочных границ в
результате возможного резкого отличия двух соседних вариантов на фоне общего
высокого сходства этой части временного ряда.
Хронологический ряд орнитокомплексов
сенокосов, чередующихся с перелесками, разделен на 6 групп, а остальных
вариантов - на 7. Дальнейшее деление приводило к вычленение в качестве
самостоятельных классов отдельных проб, поэтому признано нецелесообразным.
Оценка внутригодовой
динамики облика орнитокомплексов проведена по вышеуказанным матрицам сходства отдельно
для каждого из 6 местообитаний. Дальнейшая обработка основана на методах неметрического шкалирования. В
используемой для анализа программе объекты представляются точками на плоскости [14].
Само расположение точек не является простой проекцией из пространства большей
размерности. По используемому алгоритму близкие точки на плоскости расположены
близко, а далекие - далеко. Переход из многомерного пространства в двухмерное может
приводить к существенным искажениям во взаиморасположении объектов только в тех
случаях, когда влияние остальных, кроме первых двух, факторов весьма значимо.
Результаты фенологических наблюдений, проведенных в
районе учетных работ в те же сроки, взяты из Летописей природы Висимского государственного заповедника за 1983-84 гг.
(исполнитель фенологических разделов Л.Н. Магданова).
При описании численности и распределения птиц использована балльная оценка
А.П.Кузякина [15]. Названия птиц приведены по "Каталогу птиц СССР" [16].
РЕЗУЛЬТАТЫ
Границы сезонных аспектов. Первая
и наиболее ярко выраженная граница сезонных аспектов населения птиц во всех
обследованных местообитаниях приходится на середину апреля (рис. 1). Некоторое
усиление кочевок круглогодично встречающихся и, особенно, массовый прилет, и
пролет большой группы видов во II половине апреля обусловливают в южной тайге
Среднего Урала повсеместные и самые резкие в годовом цикле изменения облика орнитокомплексов. В это время, например, здесь появились рябинник,
певчий дрозд, белобровик, зарянка и белая трясогузка.
Часть видов, таких как зяблик, серая ворона, обыкновенная овсянка, скворец и
полевой жаворонок, начинают встречаться раньше, но во II половину апреля их
обилие сильно возрастает. Число видов, встреченных в одном местообитании во II
половину апреля, в сравнении с I-ой, повышается от 1.2 раза (с 14 до 17 видов)
- в пихтово-еловых лесах с примесью березы до 2.4 раза (с
10 до 24) - на свежих вырубках по темнохвойно-мелколиственным лесам. Увеличение
суммарного обилия птиц еще выше: от полуторакратного (с
239 до 369 особей/кв. км) - в тех же пихто-ельниках с
примесью березы, до одиннадцатикратного (с 47 до 515 особей/кв. км) - в
сенокосах, чередующихся с перелесками.
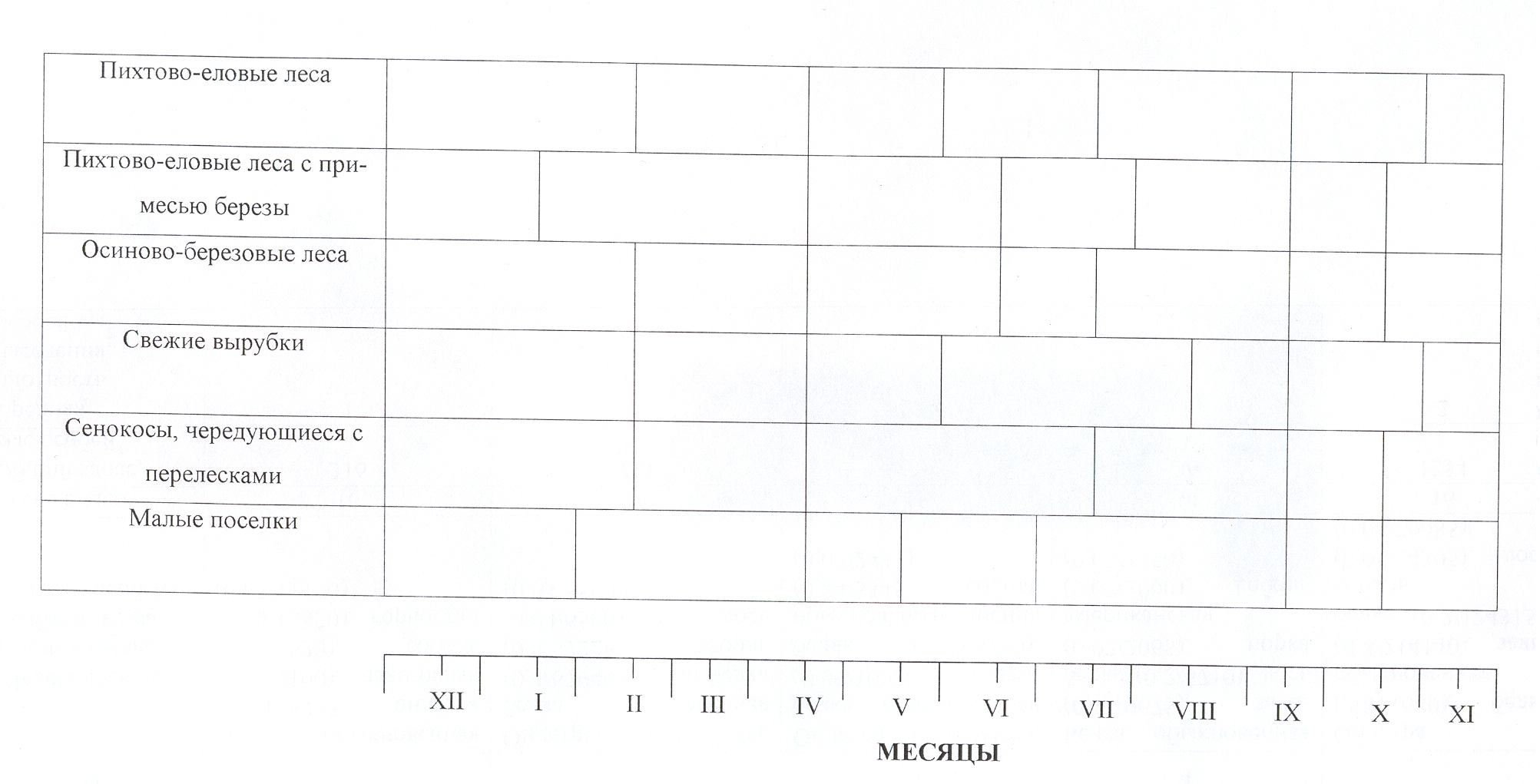
Рис. 1. Границы сезонных аспектов населения птиц южной тайги
Среднего Урала (1983-1984 гг.).
Примечание: длинные вертикальные линии в нижней части
рисунка обозначают начало и окончание месяца, короткие – их половины; римские
цифры (I – XII) – собственно месяцы.
Наиболее
значимая осенняя граница также едина для всех шести местообитаний. Она приходится
на середину сентября и определена как бы обратными по отношению к весенней
границе, причинами, т.е. снижением обилия или полным исчезновением из региона
многих видов. Начиная со II-ой половины сентября, больше не встречены
горихвостка-лысушка, зеленая пеночка, луговой и черноголовый
чеканы, деревенская ласточка. Резко снижается, в сравнении с I-ой половиной
этого месяца, обилие зяблика, теньковки, белой трясогузки и ряда других видов
(зато, например, напряженность пролета юрка и рябинника в это время выше). В
целом число встреченных в одном местообитании видов снижается повсеместно (в
1.1-2.6 раза), а плотность населения - только в лесах и на вырубках (в 1.7-3.2
раза). В сенокосах, чередующихся с перелесками, суммарное обилие, в сравнении с
I половиной сентября, почти неизменно, а в малых полузаброшенных
поселках, наоборот, почти в 2 раза возрастает.
Почти без флуктуаций
выделяется граница, вычленяющая период пролета и предзимних кочевок. Она
характеризует повсеместное снижение суммарных показателей, за счет отлета всех
перелетных видов на фоне пролета рогатого жаворонка и кочевок клеста-еловика и чечетки. В четырех местообитаниях эта
граница приходится на середину октября, в двух, смещаясь на полмесяца,
разделяет октябрь и ноябрь.
Следующая по степени общности
граница обусловлена началом предвесенних кочевок. Она тоже в четырех
местообитаниях из шести едина по срокам и приходится на середину февраля (в
пихтово-еловых лесах с примесью березы этот аспект начался в середине, а в
малых поселках – в конце января). В лесах ее проявление вызвано усилением
местных перемещений, преимущественно, пухляка и за счет этого небольшим подъемом
суммарного обилия. Начало предвесенней
активизации кочевок на вырубках и сенокосах выражено слабо и определено, в
основном, усилением перемещений тех же лесных обитателей. В малых поселках,
напротив, происходит предвесеннее (до апреля) волнообразное снижение суммарного
обилия и частичная или полная откочевка видов, обитающих в них зимой. Например,
в сравнении со II половиной января, в I половине февраля уменьшается численность
полевого воробья в 3.3, а большой синицы в 4.3 раза. Плотность населения в это
же время падает с 234 особей/кв. км до 116, а число видов с 9 до 6.
Граница, знаменующая
собой начало послегнездовых кочевок и начало отлета, значительнее
варьирует по срокам в разных местообитаниях (от конца июня в малых поселках до
середины августа на свежих вырубках). Тем не менее, в трех из шести
местообитаний она приходится на середину июля. В лесах она обусловлена, в
основном, нарастанием суммарного обилия и уходом из них некоторых видов. К
примеру, в пихтово-еловых лесах исчезают славки
- черноголовая и завирушка, малая мухоловка и т.д. Суммарное обилие возрастает в
1.5 раза. Происходит долевое перераспределение в населении массовых видов. Так, в I половине июля на долю пухляка и зяблика приходится,
соответственно, 27 и 11%, а во II - 44 и 10%. Кроме того, в I половину месяца
еще доминируют клест-еловик и зеленая пеночка (12 и
10%), обилие которых во II половину июля в 1.5-2 раза снижается, а пухляка и
зяблика увеличивается. В результате во II половину июля клест-еловик и зеленая пеночка в число доминантов не попадают.
В полуоткрытых местообитаний
на фоне нарастания суммарного обилия происходит полная смена доминантов. На вырубках
вместо лесного конька, лугового чекана и белой трясогузки начинают преобладать
теньковка, зяблик и серая славка, на сенокосах полевого жаворонка сменяют белая
трясогузка, лесной конек и серая ворона. Эта граница смены аспектов достаточно
резко отличает население птиц малых поселков от всех остальных местообитаний.
Она определяется уходом к началу июля из поселка ранее доминирующего скворца.
Одновременно, преимущественно за счет вылета молодых птиц и усилившихся
перемещений полевого воробья, деревенской ласточки и белой трясогузки,
плотность населения возрастает с 269 до 540 особей/кв.км.
Последняя из обсуждаемых границ демонстрирует наступление достаточно короткого и
значительно различающегося по срокам в разных местообитаниях периода
относительной летней стабилизации населения птиц (в сенокосах, чередующихся с
перелесками, она вообще не выражена). В лесах она связана, в основном, с
массовым появлением поздно прилетающих видов на фоне окончания пролета, поэтому
суммарное обилие меняется слабо, а общее число видов возрастает. Так, в пихтово-еловых
лесах суммарное обилие во II половину мая и I половину июня почти неизменно
(426 и 429 особей/кв. км), но, за счет прилета наиболее поздно прилетающих
видов, их общее число возрастает с 29 до 36. В массе прилетевшая и пролетающая
в начале июня зеленая пеночка в I половине июня доминирует вместе с пухляком,
сменяя преобладающего во II половине мая зяблика. На свежих вырубках
возникновение этой границы обусловлено почти 2-кратным снижением суммарного
обилия к началу июня (в сравнении со II половиной мая), вызванного окончанием
пролета у достаточно большой группы видов. К примеру, обилие белой трясогузки к
июню на вырубках снижается в 5.4 раза. В поселках появление границы вызвано
снижением во II половине мая суммарного обилия в 1.3 раза, а особенно тем, что
помимо скворца, во II половину мая начинают преобладать в населении еще
деревенская ласточка и белая трясогузка.
Проведенный анализ
внутригодовой аспективности населения птиц показал,
что сроки прохождения и значимость границ сезонных периодов могут отличаться
даже в физиономически близких местообитаниях. Так, по
сравнению с другими лесами предвесенние кочевки в пихто-ельниках
с примесью березы начинаются на месяц раньше. Эти леса состоят из более-менее
одновозрастных деревьев и местами разрежены, поэтому с удлинением к середине
января светового дня в солнечную погоду, судя по характеру и интенсивности
снеготаяния, субкоренные пихто-ельники
заметно лучше прогреваются, чем коренные. Этот тепловой вместе с прямым солнечным
воздействием может вызвать более резкую активизацию населяющих субкоренные леса птиц и вызвать дополнительный приток
особей из других местообитаний. Не хуже прогреваемые осиново-березовые леса и
вырубки в этот период очень бедны и в них подобное повышение обилия птиц не
отмечено. На свежих вырубках существенно сдвинуто
начало аспекта послегнездовых перемещенияй.
Достаточно сильное влияние на изменение облика населения здесь оказал только
самый пик кочевок и миграций, приходящийся на II половину августа. По-видимому, большая плавность изменений и
меньшая их амплитуда не позволяет выявить границу начала сезона относительной
летней стабилизации населения сенокосов, чередующихся c
перелесками. Зато этот период выражен и максимально рано наступает (и также
рано заканчивается) у населения птиц малых полузаброшенных
поселков.
Между тем, несмотря на подчас сильные
различия в составе видов и их количественном соотношении, сходство сезонной аспективности населения
птиц лесов, вырубок, сенокосов и малых поселков весьма значительно. Большинство
выявленных в разных местообитаниях периодов совпадает по вызвавшим их причинам
и близки по календарным срокам. Это позволяет выделить и условно принять для
всех местообитаний шесть общих сезонных периодов (рис. 2).

Рис. 2.
Принятые сезонные аспекты населения птиц Среднего Урала (а) в сравнении
с фенологическими сезонами года (б), 1983-1984 гг
1. Период относительной зимней
стабилизации, который длится с начала декабря до середины февраля (значимо
раньше он заканчивается только в пихто-ельниках с примесью
березы).
2. Период предвесенних кочевок и
начала прилета на 2 недели упреждает начало фенологического предвесенья
и заканчивается в середине апреля.
3. Период гнездования на фоне местных
и миграционных перемещений. Начальная граница этого периода едина для всех без
исключения местообитаний и проходит в середине апреля, на полмесяца запаздывая
по отношению к наступлению фенологической весны. Сроки его окончания менее
устойчивы, но только на свежих вырубках конец периода значимо сдвинут к осени,
а в целом заканчивается к середине июля.
4. Период послегнездовых
местных кочевок и начала отлета начинается с середины июля и приходится на
разгар полнолетья. Заканчивается этот аспект единой
границей, проходящей в середине сентября и совпадающей с окончанием глубокой осени.
5.
Период отлета и осеннего пролета длится с начала послеосенья
до середины предзимья (с середины сентября до середины октября). Флуктуации по
срокам окончания этого периода в разных местообитаниях практически незначимы.
6. Период
окончания пролета и предзимних кочевок начинается с середины октября и
захватывает II половину предзимья и часть зимы. Окончание периода приходится на
конец ноября.
Проявившийся в пяти местообитаниях из
шести период относительной летней стабилизации почти в каждом из них
значительно различался по календарным срокам и продолжительности. К примеру,
начало этого аспекта в осиново-березовых и пихтово-еловых лесах с примесью
березы лишь на полмесяца раньше его окончания в малых полузаброшенных
поселках. В целом аспект относительной летней стабилизации как бы сливается с
аспектом весенних миграций, образуя выше упомянутый (3-й) период гнездования на
фоне прилета и миграционных перемещений. Выделить общую для всех обследованных
местообитаний границу не удается, поэтому в дальнейшем описании эти периоды как
самостоятельные не рассматриваются.
Краткая количественная характеристика
сезонных аспектов.
Период относительной зимней
стабилизации отличают самые низкие в годовом цикле видовое богатство
и суммарные показатели обилия (табл. 1).
Таблица 1.
Краткая
количественная характеристика сезонных аспектов населения птиц в 6
местообитаниях Среднего Урала, 1983-84 гг.
|
Период
Местообитание
|
Относительной
зимней стабилизации
|
Предвесенних
кочевок и начала прилета
|
Гнездования
на фоне местных и миграционных перемещений
|
Послегнездовых
местных кочевок и начала отлета
|
Отлета и
осеннего пролета
|
Окончания
пролета и предзимних кочевок
|
|
Доминирующие
виды в среднем по аспекту; среднее суммарное обилие в особях/км2; всего встречено видов (из них фоновых в
среднем по аспекту)
|
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
|
Пихтово-еловые
леса
|
Пухляк, московка;
|
Пухляк, московка;
|
Пухляк, зяблик;
|
Пухляк, зяблик
московка;
|
Пухляк,
юрок;
|
Пухляк, клест-еловик, чечетка, московка;
|
|
153; 10(7)
|
218; 12(9)
|
381; 46(27)
|
677; 33(30)
|
476; 21(20)
|
271; 15(11)
|
|
Пихтово-еловые
леса с примесью березы
|
Пухляк, московка;
|
Пухляк, московка;
|
Пухляк, зяблик;
|
Пухляк, зяблик;
|
Пухляк, клест-еловик, юрок;
|
Пухляк, клест-еловик, чечетка, московка;
|
|
147; 14(8)
|
238; 19(14)
|
460; 53(36)
|
719; 33
(31)
|
480; 23(18)
|
261; 14(10)
|
|
Осиново-березовые
леса
|
Пухляк, московка;
|
Пухляк, московка;
|
Зяблик, пухляк,
юрок;
|
Зяблик,
пухляк;
|
Юрок,
пухляк, пищуха;
|
Пухляк,
чечетка;
|
|
59; 9(4)
|
142; 16(9)
|
356; 45(29)
|
552; 36(28)
|
317; 18(15)
|
157; 16(12)
|
|
Свежие
вырубки по темнохвойно-мелколиственным лесам
|
Трехпалый дятел,
малый пестрый дятел, рябчик;
|
Рябчик, обыкновенная
овсянка, пухляк, большая синица, черный дятел;
|
Лесной
конек, луговой чекан, белая трясогузка;
|
Теньковка, зяблик,
серая славка;
|
Пухляк,
чечетка, большая синица;
|
Чечетка,
рогатый жаворонок, клест-еловик;
|
|
3; 7(1)
|
11; 13(5)
|
287; 57(33)
|
320; 38(25)
|
199; 22(18)
|
71; 17(11)
|
|
Сенокосы,
чередующиеся с перелесками
|
Сорока, снегирь,
ворон;
|
Пухляк, сорока,
щегол;
|
Полевой жаворонок;
|
Белая
трясогузка, лесной конек, серая ворона;
|
Обыкновенная
овсянка, рябинник;
|
Чечетка,
рогатый жаворонок, клест-еловик;
|
|
3; 7(1)
|
22; 15(8)
|
283; 62(37)
|
343; 52(35)
|
311; 16(12)
|
197; 21(15)
|
|
Малые
поселки
|
Домовый и полевой
воробьи, большая синица
|
Домовый и полевой
воробьи, большая синица, сизый голубь;
|
Скворец,
серая ворона;
|
Белая
трясогузка, домовый воробей;
|
Полевой воробей,
рябинник, обыкновенная овсянка;
|
Чечетка,
домовый воробей, клест-еловик, щегол;
|
|
164; 12(10)
|
148; 19(11)
|
426; 57(39)
|
612; 44(31)
|
829; 24(19)
|
345; 21(16)
|
Всего
в этот период встречено 26 видов птиц. Причем, в целом за это время различные леса населяло от
9 до 14 видов, фоновых их которых в среднем было от 4 до 8 видов. На свежих
вырубках по темнохвойно-мелколиственным лесам и в сенокосах, чередующихся с
перелесками, встречено по 7 видов (фоновых - по 1), а в малых полузаброшенных поселках - 12 (10 - фоновые).
Наиболее высокое среднее суммарное обилие характерно для орнитокомплексов
малых поселков - 164 особи/кв.км, несколько ниже оно в
пихтово-еловых (153) и пихтово-еловых лесах с примесью березы (147). Плотность
населения в осиново-березовых лесах почти в 2.5-3 раза ниже - 59 особей/кв. км
и самые низкие показатели свойственны орнитокомплексам
вырубок и сенокосов (по 3 особи/кв. км). В лесах почти весь период бессменно
преобладают пухляк и московка, на долю которых в
среднем приходится 80-84%. На вырубках и сенокосах, в связи с высокой
ненаправленной изменчивостью облика населения птиц, крайне низкими суммарным
обилием и видовым богатством в отдельные отрезки времени доминировали почти все
встреченные за этот период виды. На свежих вырубках в среднем за это время преобладали
трехпалый и малый пестрый дятлы и рябчик, вместе составляя 82 % населения. В сенокосах,
чередующихся с перелесками, доминировали сорока, снегирь и ворон (41, 27 и 11
%). По стабильности облика населения птиц малые поселки близки лесным местообитаниям.
Почти весь период здесь доминировали домовый и полевой воробьи и большая синица,
составляя соответственно 31, 29 и 15 % населения.
Период предвесенних кочевок и начала прилета характеризуется усилением кочевок
зимующих видов, а конец аспекта - прилетом первых перелетных птиц. Общее число
видов возрастает до 33. Несколько богаче стал видовой состав в лесах и
поселках: в целом за период в каждом из этих местообитаний отмечено от 12 до 19
видов птиц (9-14 из которых, в среднем, фоновые).
Облик населения птиц свежих вырубок и сенокосов изменился заметнее: общее
количество встреченных видов увеличилось в 2 раза, а число фоновых в среднем по
аспекту - в 5-8 раз. В сравнении с предыдущим периодом, в лесах, на вырубках и
в сенокосах средняя плотность населения повысилась в 1.4-7.3 раза. В связи с
частичной откочевкой (особенно в начале периода) некоторых зимних синантропов
(например, большой синицы, полевого воробья) в малых поселках, наоборот, наблюдается
даже небольшое снижение этих суммарных показателей (на 8-11 %). Состав доминантов в общем сходен с предыдущим периодом. Во всех
лесах по обилию постоянно доминируют пухляк и московка.
Пухляк в среднем составлял 62-68%, а московка - 16-20
% населения птиц. В осиново-березовых лесах в начале периода в число доминантов
входит чечетка (в среднем по периоду лишь 4 %) и в конце - зяблик (в среднем –
5 %). Внутрисезонный облик населения птиц свежих
вырубок и сенокосов, чередующихся с перелесками, по-прежнему, менее стабилен,
чем в лесах и поселках. В среднем же по аспекту, на вырубках преобладают
рябчик, обыкновенная овсянка, пухляк, большая синица и черный дятел (20, 18, по
14 и 11 %). В сенокосах, чередующихся с перелесками, доминируют пухляк, сорока
и щегол (32, 13 и 11 %). Состав доминантов населения птиц малых поселков
изменился слабо. По-прежнему, доминируют воробьи и большая синица (30, 20 и 15
%), а кроме них в среднем, еще и сизый голубь (10 %).
Период гнездования на фоне местных и миграционных
перемещений включает
в себя массовый прилет и гнездование, почти все время сопровождающиеся весенним
пролетом и кочевками. В годовом цикле этот сезонный аспект характеризуется
максимальным разнообразием видов (102). В различных местообитаниях количество
встреченных за период видов возрастает до 45-62, а фоновых, в среднем по
аспекту - до 27-39. Средние суммарные показатели обилия по сравнению с
предыдущим периодом в лесах и поселках возрастают в 1.7-2.9 раза, а на вырубках
и в сенокосах - в 12.8-26 раз. За счет гораздо более резкого повышения общего
числа видов и суммарного обилия на вырубках и сенокосах в это время, в регионе
происходит значительное снижение неоднородности этих показателей. Так, в период
относительной зимней стабилизации минимальный и максимальный показатели
среднего суммарного обилия различаются в 55 раз (3-164 особей/кв. км), в период
предвесенних почевок и начала прилета в 22 раза
(11-238), а в описываемый период - лишь в 1,7 раза (279-460 особей/кв. км). В
лесах весь период доминируют пухляк и зяблик, составляя в среднем 13-31 и 13-18
% населения птиц. В осиново-березовых лесах, помимо них, в мае-июне в число
преобладающих видов входит юрок (в среднем по аспекту - 10%). В начале периода
всюду в состав доминантов входит еще и московка, а в
темнохвойных лесах - желтоголовый королек. В конце периода к постоянным
доминантам могут добавляться в различных лесах зеленая пеночка и певчий дрозд,
а в осиново-березовых - рябчик. Помимо вышеуказанных видов, в год исследований,
с конца июня в темнохвойных лесах в число доминантов входил и клест-еловик. На свежих вырубках, в сенокосах, чередующихся
с перелесками, и малых полузаброшенных поселках в
отдельные и обычно близкие временные отрезки
преобладали зяблик, белая трясогузка, рябинник и серая ворона. Однако
своеобразие облика населения и наиболее постоянных
доминантов хорошо характеризуют средние значения. Так, в среднем по аспекту на
свежих вырубках доминировали луговой чекан, лесной конек и белая трясогузка
(14, 10 и 11 %). В сенокосах, чередующихся с перелесками, преобладал полевой жаворонок
(24 %). В малых полузаброшенных поселках в это время
почти весь период доминируют скворец и полевой воробей (18 и 11 %).
Период послегнездовых
местных кочевок и начала отлета знаменует собой снижение общего числа
видов (до 79). В каждом из обследованных местообитаний общее
количество встреченных за период видов в сравнении с предыдущим периодом
также в 1.2-1.6 раза снижается, а среднее число фоновых сохраняется почти на
таком же уровне. Средние суммарные показатели обилия в это время года напротив,
достигают максимальных величин. С другой стороны отмечается увеличение пространственной
неоднородности этих показателей. К примеру, среднеминимальное
обилие (320 особей /кв. км на свежих вырубках) более,
чем вдвое ниже среднемаксимального (719 - в пихто-ельниках
с примесью березы). Состав доминантов в лесах в среднем по аспекту послегнездовых местных кочевок и начала отлета близок
предыдущему периоду. Во всех лесах по обилию и трансформируемой энергии доминируют
пухляк и зяблик. На них приходится, соответстсвенно,
по 24-28 и 13-25 % суммарного обилия. Кроме того, в пихтово-еловых лесах
преобладает московка (10 %). В среднем
по рассматриваемому аспекту в населении птиц свежих вырубок по обилию
доминирует серая славка (за счет высокого обилия в I половине периода) и
теньковка с зябликом (их обилие повышается во II половине периода), составляя,
соответственно, 10, 12 и 11 %. В сенокосах, чередующихся с перелесками,
преобладают белая трясогузка, лесной конек и серая ворона (22, 14 и 10 %).
В малых полузаброшенных поселках доминируют белая трясогузка
и домовый воробей (28 и 10 %).
В период отлета и осеннего пролета во всех 6
местообитаниях зарегистрировано 48 видов птиц. В каждом из них, в целом за
аспект, встречено по 16-34 вида. В сравнении с периодом послегнездовых
кочевок и начала отлета среднее количество фоновых видов снижается в 1.4-2.9
раза. В поселках, за счет высокой концентрации во время кочевок и пролета полевого
воробья, рябинника и обыкновенной овсянки, среднее суммарное обилие, в 1.4 раза
выше, чем в предыдущем периоде. В остальных местообитаниях эти показатели
снижаются и по значениям приближаются к таковым в период гнездования на фоне
местных и миграционных перемещений. Территориальная неоднородность суммарных
показателей обилия продолжает нарастать: в среднем по периоду они различаются в
4.2 раза. Состав многочисленных видов в разных местообитаниях в это время напротив,
становится ближе друг другу. Это обусловлено массовыми и широкими перемещениями
относительно небольшого числа видов и, в первую очередь, рябинника, пухляка, юрка,
клеста-еловика, обыкновенной овсянки и серой вороны.
В лесах и на вырубках преобладает пухляк. Вместе с ним во всех лесах доминирует
юрок. Кроме этих двух видов, в пихто-ельниках с
примесью березы преобладает клест-еловик. В
осиново-березовых лесах доминирует еще и пищуха, а на вырубках - чечетка и
большая синица. Еще более сходен список доминантов в сенокосах, чередующихся с
перелесками, и малых поселках. В обоих местообитаниях преобладают рябинник и обыкновенная
овсянка. А кроме них, в малых поселках еще и полевой воробей.
Основу
населения в период окончания пролета и
предзимних кочевок составляют птицы, зимующие в южной тайге Среднего Урала,
хотя пролет северных видов еще продолжается. Всего за этот период отмечено 36
видов птиц, а их количество в каждом местообитании за это время снижается до
14-21 вида. Среднее число фоновых видов в разных местообитаниях варьирует от 10
до 16. Происходит дальнейшее и уже повсеместное уменьшение обилия до 71-345
особей/кв. км. Эти показатели в лесах наиболее близки периоду предвесенних
кочевок и начала прилета, а на вырубках, в сенокосах и поселках могут превышать
их в несколько раз, особенно в начале периода. По доминированию видов аспект
(особенно его I половину) можно назвать периодом чечетки и клеста-еловика.
В разных местообитаниях, в среднем по аспекту, на них может приходиться по
11-33 % от суммарных значений обилия. Кроме них, во всех обследованных лесах в
это время преобладает пухляк, в среднем, соответственно, составляя 34-42 %
населения птиц. В обоих пихто-ельниках доминирует еще
московка (в среднем 11 и 13%). На свежих вырубках и в
сенокосах с перелесками, помимо чечетки и клеста-еловика,
преобладает рогатый жаворонок, в среднем составляя 16 и 18% плотности
населения. Малые поселки от всех остальных местообитаний в это время отличает
доминирующие в них, помимо клеста-еловика и чечетки,
домовый воробей и щегол (в среднем от 11 до 17% и 10 % населения).
Внутригодовая динамика населения
птиц. Годовой цикл смены облика
населения птиц подтаежных смешанных лесов Подмосковья при изображении
замкнутой линией представляет собой сильно уплощенный эллипс. В наиболее удаленных частях эллипса находятся варианты населения
летнего и зимнего периодов времени, а в наиболее близких - весеннего и осеннего
периодов [2]. На Среднем Урале летние и зимние варианты населения птиц тоже
менее сходны между собой, чем весенние с осенними.
Однако сопоставление временного сходства орнитокомплексов в разных
местообитаниях позволило выделить четыре варианта внутригодовой динамики облика
населения птиц. Первый очень близок годичному циклу смены облика
орнитокомплексов смешанных лесов Подмосковья (рис. 3). Он характерен для
населения птиц пихтово-еловых лесов (как с примесью, так и без примеси березы).
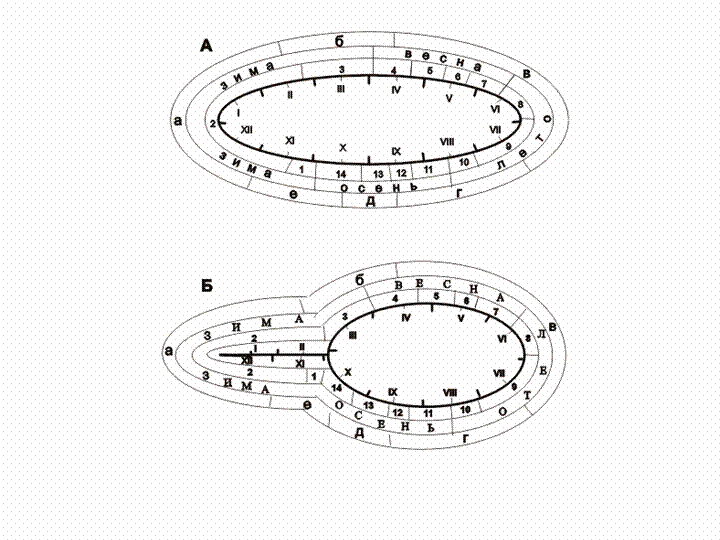
Рис. 3. Варианты сезонных циклов населения
птиц пихтово-еловых (А), осиново-березовых лесов и сенокосов, чередующихся с
перелесками (Б).
Сезонные
аспекты населения птиц: а – относительной зимней стабилизации; б - предвесенних
кочевок и начала прилета; в – гнездования на фоне местных и миграционных
перемещений; г - послегнездовые местные кочевки и начало отлета; д – отлета и осеннего
пролета; е – окончания пролета и предзимних кочевок.
Фенологические
субсезоны: 1 – первозимье; 2 – глубокая зима; 3 – предвесенье; 4 – первовесенье и
пестрая весна; 5 – оживление весны; 6 – зеленая весна; 7 – предлетье;
8 – перволетье; 9 – полное лето; 10 – спад лета; 11 –
первоосенье; 12 – глубокая осень; 13 – послеосенье; 14 – предзимье.
Второй вариант, по сути,
представляет собой эллипс и отходящую от него цепочку последовательно
отличающихся орнитокомплексов. На противоположных дугах эллипса, в его наиболее
близких частях, находятся по одной дуге варианты населения птиц позднего предвесенья и весны, по другой - осени и начала первозимья.
В наиболее удаленных частях по одну сторону - орнитокомплексы
лета с прилегающими предлетними и первоосенними
вариантами, по другую - очень близкие по облику (смыкающиеся) орнитокомплексы начала предвесенья
и окончания первозимья. От вариантов начала предвесенья
и окончания первозимья отходит цепь последовательно отличающихся
орнитокомплексов глубокой зимы (необязательно в хронологическом порядке). Такой
годичный цикл смены облика присущ населению птиц осиново-березовых лесов и
сенокосов, чередующихся с перелесками. Он связан с резким снижением в них
суммарного обилия птиц глубокой зимой и, в сравнении с первозимьем и предвесеньем, исчезновением из них ряда видов (особенно это
заметно в сенокосах). Причем, состав видов в разгар глубокой зимы продолжает
сохранять относительную стабильность, а их соотношение по обилию может меняться.
В результате зимние орнитокомплексы как бы выбиваются
из общего цикла и по мере снижения плотности населения по
нарастающей последовательно отличаются от первозимне-предвесенних
вариантов.
Годичный
цикл с крайне нестабильным населением глубокой зимой и в раннее
предвесенье свойствен свежим вырубкам по темнохвойно-мелколиственным
лесам. Он может быть изображен в виде двух примыкающих друг к другу эллипсов и
по форме кривой напоминает положенную набок цифру "8" (рис. 4).
Правая часть отражает сезонную изменчивость орнитокомплексов весны, лета, осени
и начала зимы и по соотношению вариантов весьма сходно с предыдущим типом. Его
особенности заключаются в изменчивости орнитокомплексов
зимнего периода. Эллипсовидный (а не последовательно-линейный) характер
изменчивости населения птиц глубокой зимой и в раннее предвесенье
вызывает сочетание вариабельности как суммарного обилия, в основном очень
низкого, так и крайне бедного видового состава. Образование собственно
пересечения дуг замкнутой кривой годовой динамики обусловлено значительной близостью
по облику вариантов позднего предвесенья и
предзимья-первозимья. К раннему предвесенью и началу
предзимья это сходство снижается, но в целом более высоко, чем сходство этих
орнитокомплексов с вариантами населения птиц глубокой зимы.
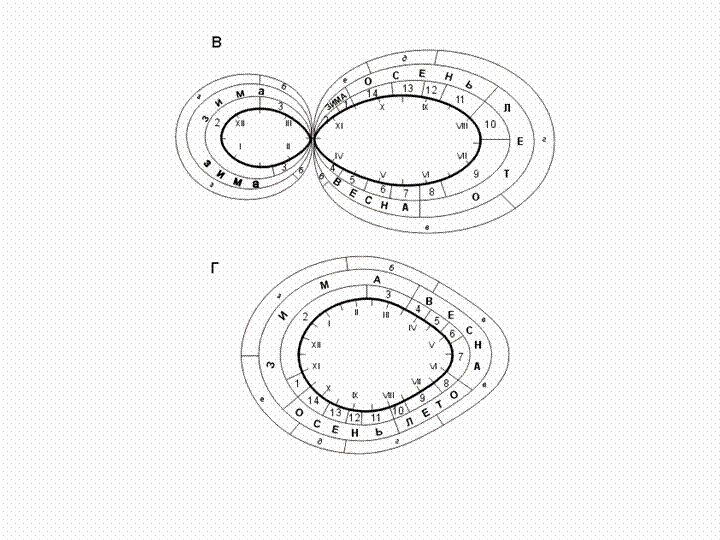
Рис. 4. Варианты сезонных циклов населения
птиц свежих вырубок (В) и малых поселков (Б).
Условные обозначения как на рис. 3.
Последний
вариант свойствен населению птиц малых полузаброшенных
поселков. Кривая динамики облика населения представляет собой
окружность с выклинивающимися орнитокомплексами
мая-июня. Изменения вариантов большей части года по окружности
обусловлены тем, что орнитокомплексы II половины лета
- первоосенья настолько отличаются от
зимне-предзимних, насколько осенние от предвесенне - ранневесенних.
Клинообразное отклонение от окружности вызвано преимущественно нарастанием в
населении птиц поселков доли скворца от начала апреля к концу июня и его
последующим исчезновением из этого местообитания к июлю.
ОБСУЖДЕНИЕ
Сравнительный
анализ собственных и опубликованных материалов показал, что по близости
внутригодовой аспективности населения птиц обследованные регионы можно объединить в две группы.
Первую составляют варианты сезонной изменчивости населения птиц юга лесной и
севера лесостепной зоны и гор юга Сибири, вторую - пустынной зоны (рис. 5).
Отличия между ними заключаются не только в разных календарных сроках смены
периодов, но и в самом характере аспективности. Так,
в южной тайге Среднего Урала, как в северной лесостепи Приобья
и на Центральном Алтае [4, 10], при обследовании территории каждые полмесяца,
разделение годового цикла населения птиц возможно лишь на шесть - семь периодов.
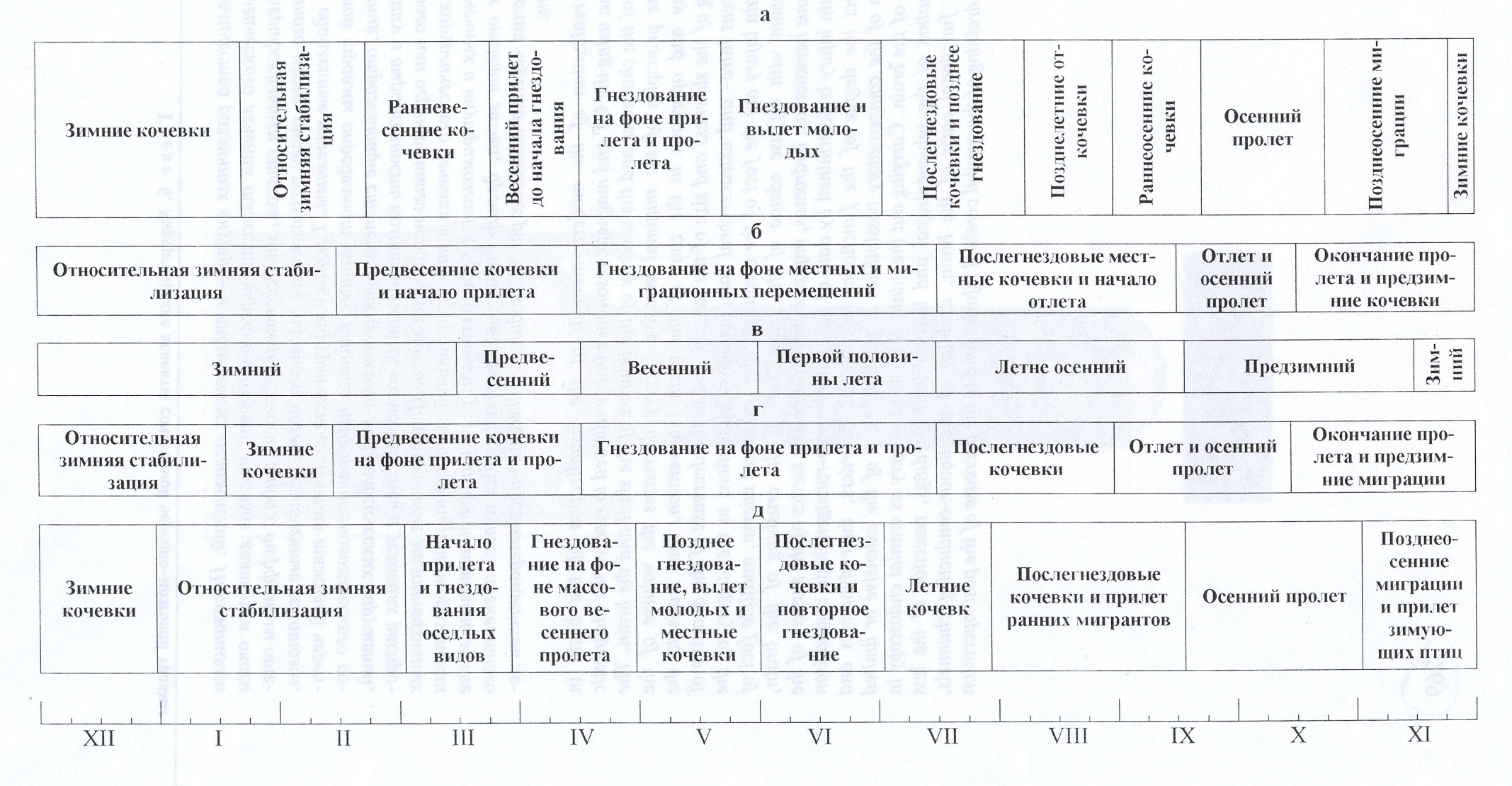
Рис. 5.
Сезонные аспекты населения птиц смешанных лесов Подмосковья (а - по Е.С. Равкину, 1985), Среднего Урала (б - по С.Г. Ливанову,
1986), северной лесостепи Приобья (в - по С.М. Цыбулину, 1985), Центрального Алтая (г – по Е.Н. Бочкаревой,
2001) и оазисов пустынь Туркмении (д
– по А.Н. Козлову, 1988).
При той же
дробности обследования, идентичных методах учета птиц и обработки полученных
данных в зоне пустыни для долин рек Мургаба и Теджена
выявлено 10 сезонных аспектов населения птиц [9]. Е.С.Равкиным
[2] для подтаежных лесов Подмосковья удалось выявить сходное количество
(одиннадцать) сезонных периодов, но при еженедельном обследовании местообитания
(т.е. при 2-кратном увеличении дробности обследования). Подобные различия в
числе выделенных сезонных периодов в целом предполагают большую внутригодовую
дифференциацию облика населения птиц в пустыне, нежели на юге бореальных
растительных областей и Алтае. Повышенная аспективность орнитокомплексов в течение года в пустынной
зоне определяется сложным сочетанием сезонной динамики местных гнездящихся птиц
и значительного количества видов, обитающих в разные отрезки внегнездовых периодов (на пролете, во время кочевок, на
зимовке). Этими же причинами объясняется и низкое межсезонное сходство
вариантов населения птиц.
Вполне понятно,
что к северу (соответственно к горам Алтая, лесостепи, подтаежным лесам и южной
тайге) число видов, населяющих эти территории только во внегнездовое
время, и их значимость в орнитокомплексах снижаются.
Иное дело, что на пролете, во время летних и зимних кочевок Алтай, север
лесостепи и юг лесной зоны в значительном количестве могут населять птицы более северных гнездовых популяций, но тех же видов, что и
местные массово гнездящиеся птицы. Подобное наложение годичных циклов
жизнедеятельности местных гнездящихся птиц и мигрантов одних и тех же видов,
вследствие их временного смещения, может даже усиливать континуальность изменений
облика населения птиц. В результате только зимние орнитокомплексы
очень резко отличаются от остальной совокупности вариантов, а в пределах
теплого или холодного периодов года характерна плавная изменчивость населения
птиц и, как следствие этого, меньшее количество сезонных аспектов.
Кроме
того, для обследованных провинций юга лесной и северной лесостепной зон и в
горах Алтая характерно хронологическое совпадение (или близость) трех-четырех границ
сезонных аспектов. Интересен тот факт, что при всех фенологических различиях и
специфике хода самой весны наиболее значимая весенняя граница аспектов в лесах
совпадает и приходится на середину апреля, почти не изменяясь в разные годы. Близость
в сроках наступления периода массового гнездования и основных волн весеннего
пролета в лесах всех вышеназванных регионов и их межгодовая устойчивость может
быть обусловлена генетической и экологической близостью населяющих эти
провинции форм. Устойчивость сроков может поддерживаться
также сочетанием толерантности к низким температурам
круглогодично обитающих и раноприлетающих птиц с
достаточной кормностью, количеством укрытий и
относительной термостатичностью лесов в это время.
В результате
облик типично лесного населения птиц относительно мало зависит от хода и
характера самой весны, что, в пределах рассмотрения, приводит к отсутствию
широтно-долготной специфики в сроках начала этого аспекта. Формирование облика населения птиц теплого периода года в полуоткрытых и
открытых местообитаниях, естественно, в значительной мере зависит от
снеготаяния и весенних снегопадов. В связи с этим начало аспекта массового
гнездования на фоне пролета, с одной стороны, может быть раньше, чем в лесах, с
другой - во многом зависит от хода весны конкретного года исследований. К
примеру, по данным С.М. Цыбулина [4] в северной
лесостепи Приобья сроки наступления этого периода в
разные годы могут варьировать в пределах месяца. Отсюда понятно, что говорить о
подзонально-провинциальной специфике наступления
весеннего аспекта населения птиц полуоткрытых и открытых местообитаний можно
лишь как о среднемноголетнем явлении. Однако по имеющимся в литературе
сведениям сделать этого невозможно. Хотя, характеризуя сезонную изменчивость населения птиц природных ландшафтов юга бореальных областей
в целом, должно отметить относительную межгодовую устойчивость весенних
сезонных аспектов (если не по календарным срокам, то по дискретности и силе
изменений облика). Подобную жесткость весенних изменений населения
безусловно определяет необходимость достаточно длительного времени для
ежегодного репродуктивного периода у птиц при сравнительно коротком оптимальном
периоде в годовом цикле развития природы. Судя по данным Е.С. Равкина [2] для смешанных лесов Подмосковья, во II половине
лета-осенью характер
сезонной аспективности населения птиц больше зависит
от фенологического хода конкретного года, вплоть до невыраженности
в отдельные годы некоторых аспектов. Даже в зимнее время, исходя из тех же
данных, в отдельные годы население птиц может отличаться по количеству
аспектов, хотя общие отличия орнитокомплексов теплого и холодного периодов
постоянно высоки.
Уральскую южную тайгу и подтаежные леса Подмосковья, от северной
лесостепи в целом отличает большее количество летне-осенних аспектов населения.
Видимо, на юге лесной зоны дискретность волн осенних миграций выражена сильнее,
чем на севере лесостепи, где птицы. Тем не менее, при общей близости границ
аспектов, приходящихся на середину сентября - ноябрь, в южнотаежных
и подтаежных лесах наблюдаются отличия в их значимости подзонального
(подзонально-провинциального) характера. Например,
наиболее значимая осенняя граница сезонных аспектов населения птиц в южнотаежных лесах Среднего Урала проходит в середине
сентября (совпадая с окончанием глубокой осени и началом предзимья). В подтаежных
лесах Подмосковья самые значительные осенние (и в годовом цикле тоже) изменения
приходятся на II половину октября, также совпадая с окончанием глубокой осени,
заканчивающейся там на месяц позже, чем в южной тайге.
Оценить
долготные отличия внутригодовой изменчивости облика населения птиц сложнее.
Вполне вероятно, что нарастание континентальности
климата усиливает асинхронность внутригодовой изменчивости облика населения
птиц по отношению к годовому циклу развития природы. На первый взгляд это
логично и отчасти подтверждается проведенными исследованиями. Действительно,
выделенные границы 11 сезонных аспектов населения птиц Подмосковных смешанных
лесов (область умеренного климата) близки среднемноголетним границам
фенологических сезонов и субсезонов [2]. С.М. Цыбулиным [4] для северной лесостепи Приобья
(область резкоконтинентального климата), напротив,
показано смещение границ аспектов населения по отношению к среднемноголетним
феносезонам. Для южной тайги Среднего Урала (область
умеренно-континентального климата) также выявлена асинхронность этих циклов.
Между тем, в лесах Подмосковья, Среднего Урала и Приобья
самая значительная из весенних границ аспектов населения и, судя по Подмосковью
и Приобью, почти не флуктуирующая по годам,
приходится на середину апреля. Она во всех трех регионах в большей или меньшей
степени, но запаздывает по отношению к началу фенологической весны. При этом в
Подмосковье эта граница приходится на начало оживления весны, а на Урале фенологически раньше - на середину первовесенья.
Самая значимая осенняя граница (и тоже, судя по Подмосковью, очень устойчивая
по календарным срокам) в Подмосковье и на Среднем Урале упреждает окончание
осени, совпадая с окончанием одного из осенних субсезонов
- глубокой осени.
Выявленные Е.С.Равкиным [2] для смешанных лесов Подмосковья границы
весенних аспектов, близкие началу и ходу развития весны, устойчивы по срокам в
разные годы, но по силе проявления в значительной мере слабее, поэтому их
выявление может быть обусловлено высокой степенью дробности проведенных исследований.
В то время как меньшая дробность учетов, выполненных на Урале и в Приобье, могла не позволить отразить эти изменения в
населении. Большинство других (поздневесенних, летних
и раннеосенних) границ аспектов
населения птиц смешанных лесов Подмосковья в разные годы могут
значительно варьировать по срокам. Из этого следует, что утверждать о наличии долготной
изменчивости в совпадении или несовпадении границ сезонных аспектов и фенопериодов, как годовых, так и среднемноголетних, видимо,
преждевременно. С уверенностью можно говорить лишь о самом характере аспективности и собственно о количестве основных аспектов
населения, которые, как показали исследования, в достаточной мере устойчивы и
закономерны. В целом, необходимо подчеркнуть нарастание количества аспектов и,
соответственно, большую дискретность изменения облика населения птиц в течение
года от бореальных растительных областей к горам юга Сибири и, особенно, к пустынной
зоне.
Для гор,
начиная с низкогорий, характерна неустойчивая весна с частыми снегопадами и
метелями. Резкие похолодания, вплоть до заморозков, возможны и в начале
фенологического лета. Такие природно-климатические особенности накладывают
отпечаток на ход динамики облика населения птиц, приводя его к ненаправленной пульсирующей
изменчивости и, как следствие, к слабой дифференциации периода относительной
летней стабилизации. На равнинах, как в Подмосковье, так и в Приобье аспект относительной летней стабилизации (именуемой
разными авторами как период гнездования и вылета молодых или период I половины
лета) выражен в населении гораздо четче.
Анализ результатов круглогодичных
учетов в малых полузаброшенных поселках южной тайги
Среднего Урала и сопоставление с литературными источниками позволяют дополнить
представления о влиянии антропогенного воздействия на годовой цикл изменчивости
населения птиц. Как показали расчеты, количество сезонных аспектов населения
птиц в этих поселках и природных местообитаниях одинаково, а их границы близки
и в большинстве своем совпадают по значимости. Так, весенняя граница аспектов,
проходящая в середине апреля едина для орнитокомплексов всех обследованных
местообитаний и наиболее значима в годовом цикле. Граница сходных аспектов
населения птиц природных и селитебных местообитаний Новосибирского
Академгородка может различаться по срокам на две недели (начало - середина
апреля), но в среднем приходится на середину апреля и наиболее значима в
сезонной периодизации облика [4]. Кроме того, как уже говорилось, количество
выделяемых для населения птиц периодов на Среднем Урале и в Приобской лесостепи
совпадает. В то же время внутригодовая изменчивость населения птиц собственно
г. Новосибирска [5] имеет свои специфические черты. Во-первых, довольно заметны
отличия в значимости границ, т.к. переход к весеннему облику населения птиц в
различных городских местообитаниях обозначен далеко не первой по значимости
границей. Во-вторых, намечается тенденция (хотя и довольно слабая) к снижению
количества сезонных аспектов: в трех из шести обследованных местообитаниях
разделения хронологического ряда вариантов населения возможно лишь на пять, а
не на шесть сезонных аспектов. Отмеченное явление подтверждается исследованиями
Е.С. Равкина [3, 17], который
на материалах учетов в московских лесопарках и подмосковных смешанных лесах
показал снижение количества сезонных аспектов в городских облесенных
местообитаниях. Постоянно достаточное количество кормов антропогенного
происхождения и благоприятный мезоклимат создают условия для большей
внутригодовой стабильности и континуальности смены облика
населения птиц крупных городов. При этом по мере нарастания степени
урбанизации, ее влияние может сказываться на характере аспективности
не только орнитокомплексов селитебных, но и природных местообитаний,
расположенных среди антропогенных ландшафтов.
ЗАКЛЮЧЕНИЕ
Итак, во всех
проанализированных регионах внутригодовая устойчивость облика орнитокомплексов снижается от населенных пунктов и лесов к полуоткрытым
и открытым местообитаниям. Внутригодовая близость облика лесных орнитокомплексов снижается от южной тайги к тугаям пустынь, поскольку на Урале обилие оседлых видов
сравнительно высоко и смена аспектов населения определяется гнездящимися
перелетными видами. В лесостепи и, особенно, в тугаях
доля видов, обитающих во внегнездовое время
(кочующих, пролетных, зимующих), значительно выше, что и обусловливает усиление
межсезонных отличий вариантов населения птиц. По характеру сезонной
изменчивости населения птиц, орнитокомплексы юга
бореальных растительных областей и пустынной зоны отличаются календарными
сроками смены периодов. Кроме того, орнитокомплексам тугаев свойственна большая внутригодовая дифференциация.
По конфигурации
годичных циклов в разных местообитаниях южной тайги Среднего Урала выделяется 4
типа внутригодовой динамики орнитокомлексов. "Эллипсовидный" - отражающий большую близость весенних вариантов
к осенним, нежели летних к зимним, при относительной стабильности последних
(пихтово-еловые леса). "Эвгленовидный" тип
представляет собой эллипс и последовательно отходящую от него цепочку зимних вариантов,
отличающихся относительно стабильным видовым составом при резком снижении
обилия птиц (осиново-березовые леса и сенокосы с перелесками). "Гантелевидный" представляет собой два примыкающих друг
другу эллипса изменений орнитокомплексов теплого периода
и зимы, где последние крайне бедны и слабо сходны между собой (свежие вырубки).
"Каплевидный" тип отражает кругообразную
изменчивость вариантов с выклиниванием орнитокомплексов
мая-июня, за счет резкого увеличения доли скворца (малые поселки).
ЛИТЕРАТУРА
1. Tomialoic L. V Konferencia Miedzynarodowego Komitetu Liczenia Ptacow oraz II Konferencia Komitetu Europejskiego Atlasu Ptacow, Szymbark k Gorlic. 15-21/IX 1976. Wiad ecol. 1977,. 23: 3, 312-314.
2. Равкин Е.С. Пространственно-временная динамика животного
населения (птицы и млекопитающие), Новосибирск, Наука, 1985, 139-159.
3. Равкин Е.С. Материалы 10 Всесоюз.
орнитол. конф. (Витебск,
17-20 сентября 1991). Ч. 1. Пленарные доклады и сообщения на симпозиумах,
Минск, 1991, 131 - 132.
4. Цыбулин С.М. Птицы диффузного города (на примере Новосибирского
Академгородка), Новосибирск, , 1985.
5. Козлов Н.А.
Птицы Новосибирска (пространственно-временная организация населения). Новосибирск,
Наука, 1988.
6. Ливанов С.Г.
Изучение птиц СССР, их охрана и рациональное использование. Тез.докл. I Съезда Всесоюз. орнитол. общ-ва и IX Всесоюз. орнитол. конф., Л., 1986, 24-25.
7. Ливанов С.Г.
Исследования природы в заповедниках Урала. Висимский заповедник:
Информационные материалы, Свердловск, 1990, 22-25.
8. Ливанов С.Г. Актуальные проблемы
изучения и охраны птиц Восточной Европы и Северной Азии. Материалы
Международной конференции (XI Орнитологическая конференция), Казань, «Матбугат
йорты», 2001, 377-378.
9. Козлов А.Н.
Птицы тугаев долин Мургаба
и Теджена и перспективы их охраны. Автореф. дис. ... канд. биол. Наук, Новосибирск, 1988.
10. Бочкарева
Е.Н. Актуальные проблемы
изучения и охраны птиц Восточной Европы и Северной Азии. Материалы
Международной конференции (XI Орнитологическая конференция), Казань, «Матбугат
йорты», 2001, 111-113.
11. Равкин
Ю.С., Ливанов С.Г., Покровская И.В. Организация научных исследований в
заповедниках и национальных парках, М., 1999, 103-142.
12. Куперштох В.Л., Трофимов В.А. Алгоритм статистической
обработки информации, Новосибирск, 1974, 88-89.
13. Наумов Р.Л.
Птицы в очагах клещевого энцефалита Красноярского края. Автореф.
дис. ... канд. биол. Наук, М., 1964.
14. Ефимов
В.М., Галактионов Ю.К., Дорощенко Ю.В. III Всесоюзная
конференция по биологической
и медицинской кибернетики, М.-Сухуми, 1978, 146-149.
15. Кузякин
А.П. Зоогеография СССР / Уч. зап.
Моск. обл. пед. ин-та. (Биогеогарфия), 1962, т.
109, вып. 1.
16. Иванов А.И.
Каталог птиц СССР, Л., Наука, 1976.
17. Равкин Е.С. Сезонная динамика населения птиц городских
лесопарков и пригородных лесов Москвы // Зоология и ландшафтная зоогеография
(чтения памяти А.П.Кузякина). М.: Наука. 1993. С. 139-157.