УДК 599.363+599.323
Пространственная
неоднородность населения мелких млекопитающих Центрального Алтая
О.П. ВОЗНИЙЧУК*, И.Н.
БОГОМОЛОВА**, С.Г. ЛИВАНОВ**, Л.Г. ВАРТАПЕТОВ**, С.В. ДОЛГОВЫХ*
*Горно-Алтайский
государственный университет
649000
Горно-Алтайск, ул. Ленкина,1
**Институт
систематики и экологии животных СО РАН
6300091
Новосибирск, ул. Фрунзе, 11
АННОТАЦИЯ
На
основании результатов многолетних учетов выявлены и описаны основные
направления изменений облика сообществ мелких млекопитающих и их иерархическая
классификация. Сопоставлены изменения населения для Северного,
Северо-Восточного, Центрального и в целом Российского Алтая.
ВВЕДЕНИЕ
Под пространственно-типологической структурой сообществ мы понимаем общий характер его территориальной неоднородности, который выявляется по взаимосвязи исследуемых вариантов населения. Пространственной организацией нами принято считать набор и взаимосвязь факторов среды, которые определяют пространственную структуру сообществ. Это организующая, упорядочивающая часть системы взаимодействия населения животных и среды [1]. Анализ пространственной неоднородности териокомплексов Центрального Алтая, ранее уже проводился [2-3]. Сбор новых данных позволил дополнить имеющиеся представления о пространственно-типологической структуре населения этой группы животных, ее структурообразующих факторах и провести оценки силы и общности связи неоднородности среды и териокомплексов.
МАТЕРИАЛЫ И МЕТОДЫ
Материал собран на хребтах Иолго, Тонгош, Семинский, Куминский, Теректинский и Катунский. Мелкие млекопитающие учитывались, как правило, с 16 июля по 31 августа в 1977-79 гг. Б.С. Юдиным, Л.И. Галкиной, А.Ф. Потапкиной [2], Ю.П. Малковым [4] и Ю.Г. Швецовым; в 1988-89 гг. С.Г. Ливановым, Л.Г. Вартапетовым; в 1996 г. С.В. Долговых, И.Н. Богомоловой и М.А. Грабовским [3, 5]; в 2000-2001 гг. О.П. Вознийчук, М.А. Лебедевой и Е.Н. Бочкаревой. Отлов насекомоядных и грызунов проводился преимущественно 50-метровыми канавками или заборчиками из полиэтиленовой пленки с пятью цилиндрами, иногда - конусами, залитыми на четверть формалином. В этом случае результаты отлова сопоставимы, поэтому показатели суммарного обилия приведены на 100 цилиндро-(конусо)-суток (ц-с). Всего проанализировано 94 варианта населения мелких млекопитающих и зарегистрирован 31 вид. Названия видов даны по Каталогу млекопитающих СССР [6], кроме арктической бурозубки (Sorex arcticus), которую, вслед за М.В. Охотиной [7], мы называем тундряной (Sorex tundrensis) и лесной мыши (Apodemus sylvaticus), которая считается самостоятельным видом [8] – малой лесной мышью (Apodemus uralensis).
Использованный нами материал собран в разные годы, различными исследователями, иногда с некоторыми отклонениями в методике, объеме и сроках. Классификация по таким данным может приводить к отклонениям отдельных проб от общих тенденций. Такие варианты могут выделяться в самостоятельные группы или попадать в несвойственные им классы, искажая закономерные отличия. С целью устранения этих недостатков мы провели усреднение данных по выделам рукописной карты экосистем Республики Алтай, составленной В.П. Седельниковым. В результате усреднения получено 19 вариантов сообществ мелких млекопитающих.
Для выявления пространственной структуры населения использован один из методов кластеризации – качественный аналог факторного анализа [9]. В качестве меры сходства использован коэффициент Жаккара [10] для количественных признаков [11]. По матрице межклассовых связей методом корреляционных плеяд [12] построен структурный граф сходства сообществ. Эта схема идеализирована, т.е. не полностью соответствует машинному разбиению. Так население пойм речных долин и таежных лиственничных лесов алгоритмически объединялись в отдельный класс, но включены нами в подтип смешанных хвойных лесов и редколесий. Такая перегруппировка соответствует нашим идеализированным представлениям о пространственной неоднородности сообществ. Сила связи пространственной дифференциации населения с факторами среды или их неразделимыми сочетаниями (природно-антропогенными режимами) с учетом перестановок рассчитана с помощью качественной линейной аппроксимации [13].
Математическая обработка материала проведена с использованием программного обеспечения банка данных лаборатории зоологического мониторинга ИСиЭЖ СО РАН. Авторы выражают глубокую признательность Л.В. Писаревской за помощь в обработке собранного материала и Ю.С. Равкину за содействие в интерпретации результатов анализа.
КЛАССИФИКАЦИЯ НАСЕЛЕНИЯ
Классификация населения мелких млекопитающих Центрального Алтая включает в себя 3 типа сообществ – гумидных, субгумидных и трансформированных. Первый объединяет население гольцов и подгольцовья, лесного и лесостепного поясов, второй – степного пояса. К третьему типу отнесено население полей и поселков.
1. Тип населения гумидных местообитаний (лидируют, %: средняя, обыкновенная и равнозубая бурозубки 20, 11 и 10, обыкновенная и красно-серая полевки по 9; плотность населения 46 особей/100 ц-с; общее число встреченных видов 28/ из них фоновых – 12; по числу особей сибирский тип фауны составляет 29% населения, европейский и тундро-лесостепные реликты по 26% (далее те же показатели приводятся простым перечислением).
Подтипы населения:
1.1 – тундр (средняя, равнозубая, тундряная и обыкновенная бурозубки 22, 21, 15 и 9, узкочерепная полевка 7; 43/18-12; сибирский тип фауны 35, транспалеаркты 26, тундро-лесостепные реликты 22);
1.2 – редколесий и смешанных хвойных лесов (средняя бурозубка 15, красно-серая и обыкновенная полевки 11 и 10, обыкновенная и тундряная бурозубки 10 и 7; 52/27-12; европейский тип фауны 31, сибирский 28, транспалеаркты 23);
1.3 – кедровых лесов (средняя бурозубка 29, красная и красно-серая полевки по 16, равнозубая бурозубка, полевка-экономка 9 и 7; 72/20-10; сибирский тип 47, транспалеаркты 36, европейский тип 8);
1.4 – мелколиственных лесов (обыкновенная и средняя бурозубки 27 и 12, темная, обыкновенная и красно-серая полевки 11, 10 и 8; 29/20-9; европейский тип 50, сибирский 21, траспалеаркты 18);
1.5 – сосновых лесов на границе с Северным Алтаем (обыкновенная бурозубка 26, обыкновенная полевка 21, средняя, тундряная и плоскочерепная бурозубки 16, 5 и 5; 38/10-10; европейский тип 63, сибирский и транспалеаркты по 16);
1.6 – лесостепи (средняя бурозубка 32, обыкновенная полевка 14, обыкновенная бурозубка 9, малая лесная и восточно-азиатская мыши 6 и 5; 30/22-8; транспалеаркты 37, европейского 32, сибирского 12).
2. Тип населения субгумидных местообитаний (обыкновенная полевка 20, малая лесная мышь 18, красно-серая полевка 16, тундряная и средняя бурозубки 10 и 9; 10/22-4; европейский тип фауны 25, сибирский 21, средиземноморский 18).
3. Тип населения трансформированных местообитаний (средняя бурозубка 23, узкочерепная полевка 21, малая лесная мышь 9, тундряная бурозубка 7 и обыкновенная полевка 6; 6/20-2; тундро-лесостепные реликты 29, транспалеаркты 28, европейский и средиземноморский типы фауны по 14).
Плотность населения в гумидном
типе почти в 5 раз выше, чем в субгумидном и в 8 раз,
чем в типе населения трансформированных местообитаний, т.е. с уменьшением
увлажнения и продуктивности биоценозов и увеличением антропогенного влияния
суммарное обилие уменьшается. Такая же тенденция прослеживается для остальных
показателей. Общее видовое богатство типа населения трансформированных
местообитаний почти не отличается от субгумидного
типа (20 и 22 вида) и примерно в 1.5 раза ниже, чем в гумидном
(28). Результаты пересчета числа видов в среднем на одно местообитание
показали, что максимальное их число приходится на гумидный
тип населения (11) и уменьшается при остепнении и антропогенной нагрузке (6
видов в степях и 5 в полях и поселках). Число фоновых видов в гумидных местообитаниях в 3 и 6 раз больше, чем в субгумидных и антропогенных.
Во всех трех типах населения есть общие лидеры –
средняя бурозубка и обыкновенная полевка, причем в гумидных
и трансформированных ландшафтных урочищах средняя бурозубка стоит на первом
месте. Красно-серая полевка в качестве лидера встречается только в гумидных и субгумидных
местообитаниях, а малая лесная мышь в субгумидных и
трансформированных. Такие виды, как обыкновенная и равнозубая
бурозубки среди лидеров отмечены лишь в гумидных
ландшафтах, а узкочерепная полевка лишь в
антропогенных.
Наибольшая доля по обилию в первом
типе населения приходится на сибирский тип фауны, во втором – на европейский, а
в третьем - на тундро-лесостепные
реликты. Второе место в гумидных сообществах
принадлежит европейскому типу, в субгумидных –
сибирскому, а в трансформированных – транспалеарктам. Третье место в трансформированных
ландшафтах поделили европейский и средиземноморский типы, в гумидных
сообществах занимают транспалеаркты, в субгумидных –
средиземноморский тип фауны. По числу видов в гумидных
местообитаниях лидирует сибирский тип
фауны, в субгумидных – европейский, в трансформированных – транспалеаркты.
Максимальная плотность населения в первом типе населения отмечена в кедровых лесах, несколько меньше – в подтипе сообществ редколесий и смешанных хвойных лесов и почти в 2.5 раза меньше в лесостепи. Общее число видов максимально в редколесьях и смешанных хвойных лесах, несколько меньше их в лесостепи и в 1.5 раза меньше в подтипах кедровых и мелколиственных лесов. Минимальное видовое богатство свойственно сосновым лесам. Фоновых видов больше всего в редколесьях и смешанных хвойных лесах. На втором месте кедровые и сосновые леса и на последнем – лесостепь. Видовое богатство и число фоновых видов уменьшается при остепнении и снижении продуктивности биоценозов.
По сравнению со средней по типу плотность населения выше в редколесьях и темнохвойных лесах, почти одинакова в тундрах и ниже в остальных подтипах. Общее число встреченных видов в редколесьях и смешанных хвойных лесах отличается на единицу от показателя по типу в целом, в лесостепи – на шесть видов. В кедровых и мелколиственных лесах значения снижаются на восемь видов, а в тундрах и сосновых лесах – соответственно на 10 и 18. Больше всего видов в пересчете на одно местообитание за сезон в подтипах приходится на лесостепь (14), причём их больше, чем в среднем по типу. Одинаковое число со средним количеством видов по типу отмечено в редколесьях и смешанных хвойных лесах (11). На один вид меньше их в тундрах и мелколиственных лесах (10) и на два – в кедровых и сосновых лесах по 9. Число фоновых видов в подтипах по сравнению с типом в целом почти не изменяется, за исключением мелколиственных лесов и лесостепи, где эти показатели снижаются на три и четыре вида.
По числу особей в Центрально-Алтайской провинции в
лесах, кроме кедровых, и
степях преобладают представители европейского типа фауны. В тундрах и кедровых
лесах – сибирского, в лесостепи – транспалеаркты, в трансформированных
ландшафтах – тундро-лесостепные реликты. По
количеству видов в большей части сообществ лидируют представители сибирского
типа фауны и только в сосновых лесах, в полях и поселках на первое место
выходит европейский тип. Последнее место везде занимают тундро-лесостепные
реликты.
В отличие от ранее опубликованной классификации [3], где в Центральном Алтае выделяется только два типа сообществ мелких млекопитающих, теперь они объединены в 3 типа. Включение в расчеты дополнительных данных позволило скорректировать классификационные представления о неоднородности населения мелких млекопитающих рассматриваемой провинции. Разделенные до этого по разным подтипам влажные и сухие тундры; мелколиственные и лиственничные лесостепи в нашей схеме объединены в единые подтипы. Выделенный ранее подтип населения субальпийских редколесий в сочетании с лугами по дополнительным данным объединен с сообществом смешанных хвойных лесов. Сообщества мелколиственных лесов в отличие от предыдущей классификации образовали отдельный подтип. В тоже время, выявленная ранее пространственная неоднородность основных суммарных показателей подтвердилась. Плотность населения по выделенным типам, как и прежде, уменьшается при остепнении, уменьшении влагообеспеченности и увеличении антропогенного влияния. Видовое богатство, как и число фоновых видов, в гумидном типе также выше, чем в субгумидном. Как и в предыдущей классификации в подтипах гумидных сообществ больше всего суммарное обилие в редколесьях, темнохвойных и смешанных лесах и тундрах, меньше в сосновых лесах и лесостепи. В типах обеих классификаций есть общие лидеры – средняя бурозубка и обыкновенная полевка, наибольшая доля по обилию приходится на сибирский тип фауны в гумидных и на европейский – в субгумидных типах. По фаунистическому составу также лидируют сибирские, европейские виды и транспалеаркты.
Таким образом, при классификации населения мелких млекопитающих Центрального Алтая прослеживается влияние увлажнения, теплообеспеченности, продуктивности, антропогенной трансформации ландшафтов и, в меньшей степени, состава лесообразующих пород, остепнения и абсолютных высот местности.
ПРОСТРАНСТВЕННО-ТИПОЛОГИЧЕСКАЯ
СТРУКТУРА НАСЕЛЕНИЯ
При пороге значимости сходства в 36 единиц часть из восьми выделенных при классификации подтипов населения мелких млекопитающих образует вертикальный ряд (1-2, 6-7), который иллюстрирует изменения, связанные с влиянием поясности, абсолютных высот местности, степени облесенности и увлажнения (см. рис.). Воздействие этих факторов скоррелировано, поскольку с увеличением абсолютных высот теплообеспеченность уменьшается, а увлажнение возрастает. Сочетание тепла и влаги (гидротермический режим) определяет поясность растительности. Отклонение от основного тренда связано с различиями в составе лесообразующих пород (3-5 подтипы) и антропогенным влиянием (8).
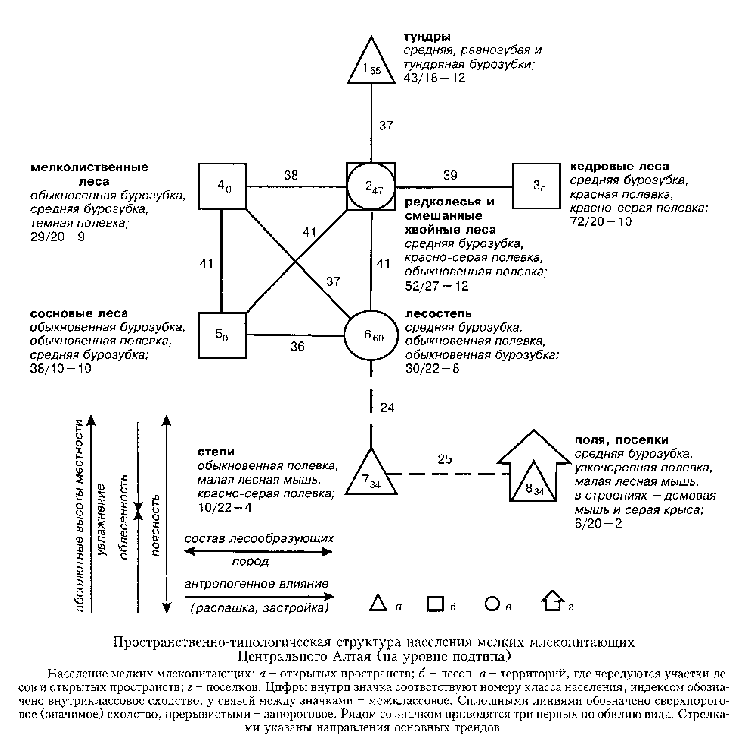
Пространственно-типологическая структура населения мелких
млекопитающих Центрального Алтая (на уровне подтипа)
Условные обозначения: население мелких млекопитающих:
а – открытых пространств; б – лесов; в – территорий, где чередуются участки
лесов и открытых пространств; г – поселков. Цифры внутри значка соответствуют номеру класса населения,
индексом обозначено внутриклассовое сходство,
у связей между значками – межклассовое. Сплошными линиями обозначено
сверхпороговое (значимое) сходство, прерывистыми – запороговое.
Рядом со значком приводиться три первых по обилию вида. Стрелками указаны
направления основных трендов.
Наибольшее суммарное обилие мелких млекопитающих в основном ряду
схемы свойственно редколесьям и
смешанным хвойным лесам с оптимальным гидротермическим режимом. Оно
незначительно снижается с уменьшением теплообеспеченности
и облесенности в тундрах (в 1.2 раза) и с уменьшением абсолютных высот,
влажности и увеличением теплообеспеченности – через
лесостепь к степям (в 2 и 5 раз). Видовое богатство изменяется так же, но без
отличий между лесостепью и степями. Число фоновых видов увеличивается в
соответствии с абсолютными высотами местности. Отклонение от основного тренда,
связанное с изменением состава лесообразующих пород, характеризуется
увеличением по сравнению с редколесьями и смешанными хвойными лесами плотности
населения на треть в кедровых лесах и уменьшением на четверть в сосновых и
вдвое – в мелколиственных лесах. Распашка и застройка не вносят значительных
изменений в показатели видового богатства, но заметно снижают
число фоновых видов и общее обилие зверьков.
Таким образом, анализ структурного графа показывает, что пространственные
тренды в населении мелких млекопитающих Центрального Алтая коррелируют
в основном с поясным типом растительности. Отклонения от основного тренда
определяются различием в составе лесообразующих пород и антропогенным влиянием.
Добавление сравнительно небольших объемов нового материала внесли некоторые коррективы в ранее составленную схему пространственно-типологической структуры населения мелких млекопитающих Центрального Алтая [3], однако выявленные тренды обеих структур имеют общую направленность, связанную с поясностью, увлажнением, облесенностью и составом лесообразующих пород.
Основные направления изменений сообществ мелких млекопитающих Северо-Восточного Алтая также связаны с абсолютными высотами местности и поясностью [14]. При этом на обобщенной схеме выделяется население предгорной лесостепи; предгорных лесов; низкогорно-среднегорных лесов нормальной полноты; тундр и редколесий; а также разреженных лесов низкогорий. Максимальное обилие млекопитающих в этой провинции свойственно черневому низкогорью и, видимо, в отдельные годы среднегорным субальпийским редколесьям с высокотравными лугами. Население среднегорных темнохвойных таежных лесов гораздо беднее (за исключением лет с урожаем семян кедра). Для Северного Алтая увеличение абсолютных высот местности также определяет пространственные изменения населения мелких млекопитающих. Кроме того, прослеживается влияние экспозиции склонов: в среднегорных лесостепных ландшафтах на склонах южной и юго-западной экспозиций население зачастую похоже на сообщества низкогорных лесов по северным и северо-восточным склонам. При увеличении остепнения на южных и юго-западных склонах того же уровня и по направлению вверх и вниз происходит обеднение населения, при этом сокращается обилие бурозубок и возрастает долевое участие полевок [15]. В этой провинции плотность населения зверьков максимальна в лесных среднегорных ландшафтах.
Итак, в Северо-Восточном, Северном и Центральном Алтае прослеживается значительное сходство в изменении сообществ мелких млекопитающих в зависимости от высоты местности над уровнем моря. По выше упомянутым провинциям и Юго-Восточному Алтаю выявлена обобщенная пространственная структура [16]. Ее анализ показывает, что неоднородность териокомплексов на этой территории, так же как по отдельным провинциям, коррелирует с абсолютными высотами местности и определяется сменой растительности. При этом можно говорить о четырех значимых для мелких млекопитающих градациях тепло- и влагообеспеченности, то есть делить территорию на теплые, умеренно теплые, умеренно холодные и холодные местообитания, а также на влажные, умеренно влажные, умеренно сухие и сухие. Сочетанием этих градаций и определяется структура населения. Кроме общих с Центральным Алтаем трендов, связанных с абсолютными высотами местности и влагообеспеченностью для Российского Алтая, судя по обследованным провинциям, проявляются отличия, определяемые провинциальностью, а также наличием или отсутствием снежного покрова и его глубиной. Общая тенденция изменения плотности населения и видового богатства вдоль основных трендов совпадает с таковой в Центральном Алтае.
Таким образом, пространственные структуры населения Северо-Восточного, Северного, Центрального и в целом Российского Алтая похожи по направленности территориальных изменений населения мелких млекопитающих в факторном пространстве и имеют отличия, связанные с высотно-поясной и экспозиционной сменой ландшафтов и провинциальными различими. Так, Северо-Восточный Алтай отличается наибольшей влажностью, мощными почвами и густотой травянистого яруса, что обеспечивает благоприятные условия существования для многих видов зверьков. Центральный Алтай суше предыдущих провинций, мощность почв здесь намного меньше, снежный покров распределен неравномерно, а в степях часто отсутствует. В связи с этим плотность населения мелких млекопитающих в Центральном Алтае почти в четыре и шесть раз ниже суммарного обилия в Северной и Северо-Восточной провинциях.
ПРОСТРАНСТВЕННАЯ ОРГАНИЗАЦИЯ НАСЕЛЕНИЯ
Оценка силы и общности связи неоднородности среды и животного населения проведена двумя способами – по данным, усредненным по выделам карты экосистем, и без усреднений (по ландшафтным урочищам). Для сравнения приведены ранее рассчитанные оценки [3]. Всего при классификации и построении структурного графа выявлено 8 факторов среды (см. табл.).
При индивидуальной оценке силы связи изменчивости населения мелких млекопитающих и факторов среды в обоих вариантах наиболее значимой является высотная поясность растительности. Этим фактором, в зависимости от варианта расчетов, объясняется 64 и 22 % дисперсии матрицы. Заметим, что высокий процент учтенной дисперсии при усреднении обусловлен нивелированием пространственных и временных отличий, а также незначительным размером матрицы. Второе место среди факторов по усредненным данным занимает состав лесообразующих пород, тогда как по исходным данным вместе с увлажнением он занимает только третье место. Не менее значима сила и общность связи неоднородности населения и облесенности. Увлажнение в первом варианте, в отличие от второго, занимает лишь четвертое место и снимает 34% дисперсии. С антропогенным влиянием можно связать лишь 29 и 7%. Степень совпадения неоднородности населения и абсолютных высот невысока и занимает предпоследние места, еще меньше влияние закустаренности.
Множественная оценка связи всех выявленных факторов по усредненным данным составляет 86% дисперсии матрицы коэффициентов сходства, по исходным – 33%. Природными режимами по классификации можно объяснить соответственно 77 и 18%, а по структуре – 70 и 17% дисперсии. Всего в обоих случаях факторами и режимами учитывается 90% дисперсии (коэффициент регрессии 0.94) и соответственно по исходным данным – 35% (0.59).
Анализ результатов расчета по ранее опубликованным данным и дополнительной выборке по всем ландшафтным урочищам за некоторым исключением совпадают. Увлажнение по нашим результатам менее значимо, чем зональность и облесенность, в отличие от сравниваемых данных, где увлажнение занимает второе место. В дополнительно собранных материалах переувлажненные урочища, не характерные для Катунского хребта, не представлены, поэтому оценки по этому фактору становятся менее значимыми. Общее совпадение результатов говорит о достаточности материалов и надежности описания структуры и классификации.
ЗАКЛЮЧЕНИЕ
Таким образом, в населении мелких млекопитающих можно выделить три типа сообществ – гумидных, субгумидных и трансформированных, причем показатели характеризующие сообщества первого типа выше таковых второго и третьего. Пространственная неоднородность населения мелких млекопитающих центральной части Алтая определяется в первую очередь поясным типом растительности, абсолютными высотами местности, степенью облесенности и в меньшей степени различием в лесообразующих породах, а также антропогенным влиянием. Пространственные тренды населения Северо-Восточного, Северного, Центрального и в целом Российского Алтая имеют общую направленность и отличаются в связи с провинциальными особенностями. Уменьшение увлажнения и смена ландшафтов от таежных и лесостепных до мозаичных и открытых среднегорно-высокогорных в ряду Северо-Восточный – Северный – Центральный Алтай, приводит к снижению суммарного обилия мелких млекопитающих в этих провинциях (89; 60 и 16 особи/100 ц-с). Оценка силы и общности связи факторов среды и неоднородности населения показывает, что наиболее значимы из них высотная поясность, состав лесообразующих пород, облесенность и увлажнение. Существенно меньше совпадение изменения с абсолютными высотами и закустаренностью. Информативность представлений о структурообразующих факторах среды и природно-антропогенных режимах по усредненным данным составила 90% учтенной дисперсии коэффициетнов сходства териокомплексов, что говорит о достаточной полноте набора выявленных факторов и режимов.
ЛИТЕРАТУРА
1.
Ю.С. Равкин, Птицы лесной
зоны Приобья, Новосибирск, Наука, Сиб.
отд-ние, 1978.
- Б.С. Юдин, Л.И. Галкина, А.Ф. Потапкина,
Млекопитающие Алтае-Саянской
горной страны, Новосибирск, Наука, Сиб. отд-ние, 1979.
- С.В. Долговых, И.Н. Богомолова, С.Г. Ливанов и
др., Сиб. экол.
журн., 1999, 6: 5, 573-584.
- Ю.П. Малков, Н.П. Малков, С.М. Цыбулин и др., Вопросы экологии и охраны позвоночных
животных, вып. II: сб. науч.
тр., Киев-Львов, Украинская академия медицинских
и биологических наук, 1998, 93-110.
- С.В. Долговых, И.Н. Богомолова, С.Г. Ливанов и
др., Доклады на международном симпозиуме «Модели устойчивого
социально-экономического развития Республики Алтай и стран Алтай-Саянского региона»,
Горно-Алтайск, 1997, 57-79.
- Каталог млекопитающих СССР. М., 1981.
- М.В. Охотина, Зоол.
журн., 1983, 62: 3, 409-417.
- И.М. Громов, М.А. Ербаева,
Млекопитающие фауны России и сопредельных регионов. Зайцеобразные и
грызуны. С.-П., 1995.
- В.А. Трофимов, Ю.С. Равкин,
Количественные методы в экологии животных. Л., 1980, 113-115.
- P. Jaccard, Bull. Soc. Vaund. Sci. Nat., 1902, 38, 69-130.
- Р. Л. Наумов, Птицы природного очага клещевого
энцефалита Красноярского края, Автореф. дис. … канд. биол. наук, М., 1964.
- П.В.
Терентьев, Вестн. Ленингр. ун-та. Сер. биол., 1959, 9, 137-141.
- Ю.С. Равкин, В.Л. Куперштох, В.А. Трофимов, Методика выявления
пространственной организации населения птиц в диалоговом режиме
«специалист - ЭВМ», Всесоюз. орнитолог. конфер.,
Киев, 1977. ч. I. с.
24-25.
- В.И. Шадрина, Фауна и экология позвоночных
Сибири, Новосибирск, Наука, Сиб. отд-ние, 1980, 79-88.
- С.М. Цыбулин, Н.И.
Богомолова, Пространственно-временная динамика животного населения,
Новосибирск, Наука, Сиб. отд-ние,
1985, 188-197.
- Ю.С. Равкин, С.М. Цыбулин, С.Г. Ливанов и др. Биоразнообразие
и динамика экосистем Северной Евразии: информационные технологии и
моделирование (WITA`
2001). ИЦиГ, Новосибирск, 2001. с.73.