УДК 597.6
ПРОСТРАНСТВЕННО-ТИПОЛОГИЧЕСКАЯ
СТРУКТУРА НАСЕЛЕНИЯ ЗЕМНОВОДНЫХ ЗАПАДНО-СИБИРСКОЙ РАВНИНЫ
Ю.С.Равкин, Л.Г.Вартапетов,
В.А.Юдкин, И.В.Покровская, И.Н.Богомолова, С.М.Цыбулин, В.Н.Блинов, В.С.Жуков,
А.К.Добротворский, Т.К.Блинова, В.П.Стариков, В.М.Ануфриев, К.В.Торопов,
С.А.Соловьёв, Г.М.Тертицкий, Е.Л.Шор
Аннотация
На основании учётов, проведённых 50-метровыми
канавками и заборчиками с пятью ловчими цилиндрами, проанализирована
неоднородность населения земноводных всех ландшафтных зон Западной Сибири,
описаны широтные изменения плотности населения и видового богатства в целом по
зонам, подзонам и группам ландшафтов, выявлена пространственно-типологическая
структура сообществ.
Под
пространственно-типологической структурой животного населения мы понимаем общий
характер его территориальной неоднородности, которая определяется
(организуется) факторами среды и взаимоотношениями жи-вотных [1]. Её выявление
для сообществ земноводных осложняется коле-баниями их численности по годам,
разной удалённостью ловчих канавок от водоёмов выплода и локальным влиянием
увлажнения, т.е. тем, что уровень их обилия не всегда определяется ландшафтными
условиями среды в целом. Тем не менее, ландшафтное урочище принято нами в
качестве наименьшей территориальной единицы рассмотрения для получения
сопоставимых данных по разным группам исследуемых животных (земноводные, птицы,
мелкие млекопитающие). Поэтому выявленные особенности населения амфибий
соответствуют результатам анализа в мелком и среднем масштабе рассмотрения.
Материалы и методы
Материалы по численности
земноводных, собранные в разные годы, но во многих местах, в полных ландшафтных
аналогах или в пределах сходных ландшафтов и групп местообитаний,
представительны не только в пространстве, но и во времени. Однако для надёжного
выявления территориальных отличий требуются дополнительные усилия, нивелирующие
временную изменчивость. В частности, такие данные можно сначала усреднить,
перейдя в более крупный ранг ландшафтного деления, скажем, с ландшафтных урочищ
на местности или ландшафты. При этом, естественно, часть информации о
внутриландшафтной неоднородности сообществ будет утеряна, но отличия более
крупного ранга (на уровне групп ландшафтов, зон, подзон, провинций), а также
различия населения ландшафтов по градиентам основных структурообразующих
факторов среды будут проявляться чётче, т.к. годовые отличия в результате
усреднения будут снивелированы. Однако такое усреднение на ландшафтной или
геоботанической основе может привести к существенным потерям информации, если
внутри ландшафтов имеются значимые отличия в населении и сообщества части
урочищ одного ландшафта более похожи на население урочищ другого ландшафта.
Кроме того, в больших выборках неизбежно накапливаются пробы, аномально отличающиеся
от всей совокупности случайно из-за дефекта сбора (погоды, объёма материала)
или необычного окружения. Эти пробы затрудняют не только интерпретацию
различий, но и анализ групп, представительных по числу проб, поскольку при
формализованном разбиении они нередко объединяются исключительно потому, что
имеют, в отличие от аномалий, хоть какое-то сходство значимого уровня. Поэтому
обширные материалы из-за перечисленных особенностей приходится сначала
разбивать на группы тем или иным способом для уменьшения объёма расчётов и проводить
усреднения для нивелировки временных отличий.
В данном случае для этой
цели сначала была проведена автоматическая классификация населения [2]
раздельно по каждой из зон, а в лесной зоне - подзон. В качестве меры
сходства-различия использован коэффициент Жак-кара-Наумова. Затем по
результатам классификации создавались концептуальные классификационные схемы,
отражающие общие особенности неоднородности сообществ. После этого данные
усреднены по сформированным классам и на этих средних выявлена структура населения
земноводных Западно-Сибирской равнины в целом. Такой подход сопровождается меньшими
потерями информации, поскольку в классы объединяются, как правило, похожие
варианты населения. Правда, причины их сходства могут быть весьма различны и в
отдельных случаях в группы могут объединяться непохожие между собой варианты,
если они по составленной классификационной схеме одинаковы концептуально
(типологически). Фактически их различия могут быть значительными, но объяснить
их в выбранном масштабе и ранге, в котором собирался материал, не удаётся.
Для анализа изменений
основных показателей сообществ земноводных по градиенту зональности и внутри
зон материалы предварительно усреднены по зонам (в лесной зоне по подзонам)
дифференцированно по группам ландшафтов: суходольные местообитания (без
населённых пунктов), внепойменные болота, поймы крупных рек, посёлки. Расчёты
средних проведены по соотношению площадей, занимаемых отдельными
местообитаниями. Их площади замерены по карте “Растительность Западно-Сибирской
равнины” (1976), без поправок на проекционные искажения, которые составляют примерно
+2%.
Материалы, послужившие
основой для настоящего сообщения, собраны вкладчиками банка данных лаборатории
зоологического мониторинга Института систематики и экологии животных СО РАН в
1970-1991 гг., как правило, за период с 16 июля по 31 августа [3]. Основная их
часть характеризует население земноводных только в один из летних сезонов, лишь
на некоторых ключевых участках учёты проводились в течение ряда лет. Всего,
считая с многолетними и литературными данными, использованы результаты
обследования 667 вариантов населения земноводных. Часть сведений опубликована
ранее [4-19].
Большая часть материалов
собрана методом отлова земноводных в 50-меторовые канавки или заборчики из полиэтиленовой
плёнки с 5 цилиндрами. Все показатели обилия даны в пересчёте на 100
цилиндро-суток (ц/с).
Градиентные изменения населения
Западно-Сибирская равнина
заселена земноводными от южной субарктической тундры до степи включительно, но
лишь остромордая лягушка распространена на всей этой территории. Серая жаба и
сибирский углозуб прослежены нами только от северной тайги до лесостепи, хотя
последнего отмечали и севернее - вплоть до южной субарктической тундры [20].
Сибирская лягушка попадалась нам от средней тайги до лесостепи, но встречается,
судя по той же публикации, и южнее. Обыкновенный тритон занимает, в основном,
лесостепь и подтаёжные леса, хотя отдельные находки имеются в южной и средней
тайге. Обыкновенная чесночница встречена в тоболо-ишимских подтаёжных лесах и
лесостепи, а также в степи Тургайской депрессии. Зелёную жабу отлавливали в
степной зоне на западе равнины. Кроме того, её изолированная популяция
обнаружена на окраине Новосибирска, расположенного в лесостепной зоне [21].
Озёрные лягушки нередко встречаются в пределах лесостепи на Оби и в Обском
водохранилище, а также в Томске (южная тайга). Видимо, они были неоднократно
завезены при рыборазведении или для лабораторных целей и случайно выпущены
[22].
Таким образом, в арктической
тундре, а также в северной и средней полосах субарктических тундр земноводные
не обнаружены. В южной тундре и лесотундре нами встречен только один вид
амфибий, в северной тайге - три, в средней - четыре, в южной - пять, в
подтаёжных лесах - шесть, в лесостепи - восемь видов, причём два из них
завезены человеком. В степи отмечено три вида земноводных. Следовательно, их
видовое богатство убывает к северу и к югу от лесостепи.
Плотность населения
земноводных, как пойкилотермных животных, напрямую зависит от общей
теплообеспеченности территории и поэтому уменьшается к северу. Их обилие
зависит также от кормности угодий (продуктивности биоценозов) и увлажнения,
следовательно, плотность населения должна убывать к югу и северу от оптимальной
гидротермической и максимальной по реальной продуктивности широтной полосы
(южной тайги или подтаёжных лесов). Однако, в действительности, судя по нашей
выборке, изменение плотности гораздо сложнее (табл. 1). Так, на суходолах максимальное
суммарное обилие свойственно средней тайге и степи. В первом случае это
объясняется высоким обилием серой жабы, во втором – обыкновенной чесночницы и,
отчасти, зелёной жабы. В степи увеличение отмечается только в западной части
равнины, поскольку два последних вида не встречаются в восточной ее половине.
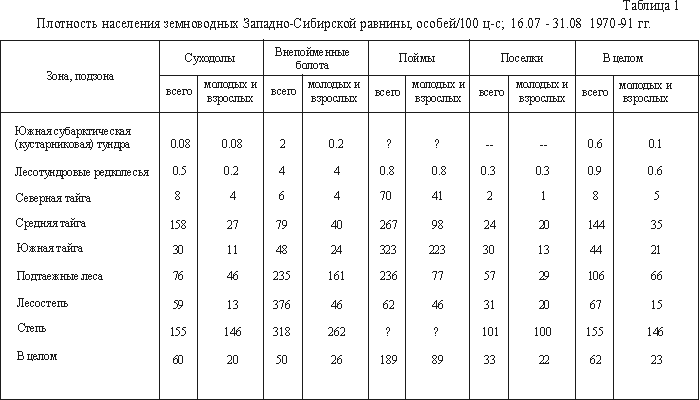
На внепойменных болотах, где
земноводные не испытывают отрицательного влияния аридизации и одновременно
пользуются всеми благами высокой топлообеспеченности, их больше всего в
лесостепи, а без учета сеголеток - в степи. К северу суммарное обилие убывает,
но на средне-таежных болотах, так же, как на суходолах, отмечено увеличение
плотности населения по сравнению с южной тайгой. Последнее свойственно, в
основном, верховым болотам, где в средней тайге вдвое больше серой жабы. В
южной тайге такие болота значительно беднее земноводными из-за большей
олиготрофности и, соответственно, меньшей кормности, а также в связи с высокой
кислотностью водоемов.
В поймах наибольшее
количество амфибий характерно для южной тайги и далее неуклонно убывает к северу
и югу. В поселках максимальная плотность зарегистрирована в степи - за счет
зеленой жабы. Следующими по уровню обилия идут поселки подтаежных лесов, хотя
земноводных здесь почти вдвое меньше. К северу от них и южнее (в лесостепи)
обилие земноводных уменьшается.
В целом по зонам и подзонам
максимальная плотность населения свойственна степи, затем значения убывают к
северу, хотя в подтаежных лесах и, в средней тайге неуклонность уменьшения
нарушается. Причем, это прослеживается не только по общему обилию, но и по
показателям, рассчитанным лишь по молодым и взрослым особям. Их численность
меньше подвержена годовым колебаниям, чем сеголеток, обилие которых существенно
зависит от успешности выплода в данном году.
С севера до подтаежных лесов
внутри зон и подзон (кроме лесотундры) максимальные значения свойственны поймам
крупных рек. В подтаежных лесах, лесостепи и степи самые высокие показатели
отмечены для вне-пойменных болот. Это связано с тем, что в них поймы
сравнительно сухие, а болота почти исключительно евтрофные. К северу, начиная с
южной тайги, преобладают по площади олиготрофные болота, а с северной тайги – гетеротрофные.
В лесотундре, так же, как и на юге, максимальное обилие свойственно болотным
ландшафтам. В целом по равнине больше всего земноводных (в пересчете на 100
ц/с) держится в поймах, в 3-3,5 раза меньше их на незастроенных суходолах и
внепойменных болотах, и почти в шесть раз меньше в поселках.
В тундре и лесотундре
отмечена только остромордая лягушка. В се-верной тайге к ней добавляются сибирский
углозуб и серая жаба, на долю которых (преимущественно углозуба) приходится
всего 2% населения земноводных (табл. 2).
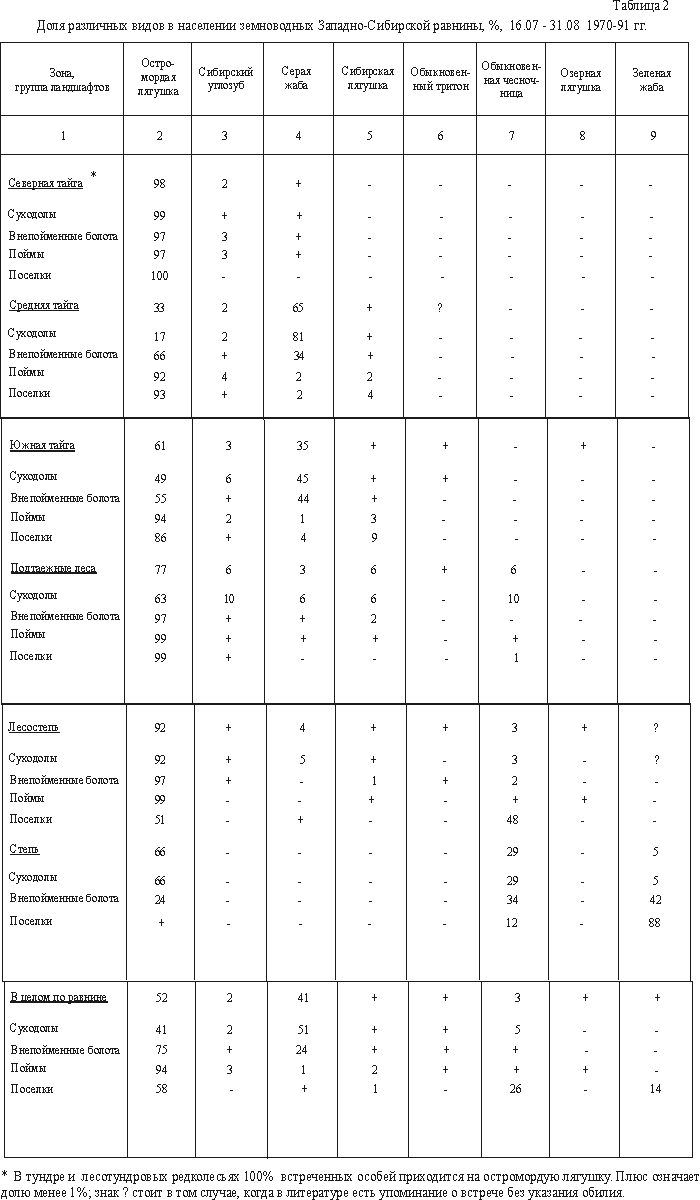
В средней тайге остромордая лягушка составляет
примерно третью часть населения, доля серой жабы вдвое больше, остальное приходится,
в основном, на сибирского углозуба. В южной тайге соотношение остромордой
лягушки и серой жабы противоположно отмеченному в средней тайге, а доля
углозуба почти не меняется. Участие сибирской лягушки значимо только в поймах.
В подтаежных лесах тоже больше всего остромордой лягушки (77%), по 6%
приходится на сибирскую лягушку и обыкновенную чесночницу, вдвое меньше - на
углозуба, а на обыкновенного тритона менее 1%. В лесостепи доля остромордой
лягушки возрастает до 92%, серой жабы и обыкновенной чесночницы равна 3 и 4%,
остальных - менее 1%. В степной зоне участие остромордой лягушки снижается до
66%, чесночницы увеличивается до 29%, а зеленой жабы составляет 5%.
На Западно-Сибирской
равнине, кроме ее юго-западной части, плотность населения фактически связана с
распределением остромордой лягушки и серой жабы. Максимальное обилие первого
вида в целом по зонам, а в лесной зоне по подзонам приходится на подтаежные
леса. К северу и югу от этой подзоны оно убывает почти неуклонно с одним
единственным исключением в средней тайге. В последней отмечен максимум обилия
серой жабы, хотя в северной тайге ее обилие резко снижается, а начиная с лесотундры,
она вообще не встречена. К югу обилие этой жабы снижается более плавно, но все
же резче, чем остромордой лягушки, и в степной зоне она нами не обнаружена.
Максимальные значения внутри зон и подзон для остромордой лягушки свойственны
лесостепным болотам, а для серой жабы - среднетаежным суходолам. Некоторое
увеличение здесь отмечено и у первого вида. Причина этого всплеска в средней
тайге скорее всего в локальных отличиях по увлажнению, связанных с соседством и
чередованием относительно узких полос сосняков и переувлажненных местообитаний.
Это создает благоприятные условия для использования первых серой жабой,
поскольку она периодически может восполнять влагопотерю в мелких водоемах и
переживать сухие дни на близлежащих болотах. Аналогичное увеличение численности
серой жабы в сосняках, граничащих с заболоченными участками отмечено в
Белоруссии [23]. Остромордая лягушка быстрее теряет влагу и не может столь
эффективно использовать эти сосняки из-за их сухости. Такое же чередование
местообитаний есть и в северной тайге, но суровость ее климата создает в целом
неблагоприятные для земноводных условия.
Возможно, часть наших сборов пришлась на депрессию численности серой жабы, другая, особенно в западной и восточной части средней тайги, - на период ее высокой численности, т.е. различия обусловлены годовыми колебаниями обилия земноводных. Видимо, для них, в отличие от мелких млекопитающих, характерны более плавные увеличения и спады обилия, причем не синхронные у всех видов. Так, в южной тайге Приобья на меж-дуречьях в 1990 г. отмечено увеличение численности серой жабы в 153 раза по сравнению с 1970 г. и десятикратное уменьшение обилия остромордой лягушки, а в низкой прирусловой пойме оби, где серой жабы нет, обилие остромордой лягушки увеличилось на 20%. В южной тайге Прииртышья среднее обилие серой жабы в 1991 г. возросло по сравнению с 1971 г. втрое, а остромордой лягушки уменьшилось в 8 раз. В то же время в лесостепных борах близ Новосибирска отмечено незначительное увеличение численности остромордой лягушки при заметном увеличении обилия серой жабы. Не исключено, что они в какой-то мере конкурируют и остромордая лягушка, как более подвижное животное, конкурентноспособнее на суше, а серая жаба, видимо, угнетает развитие головастиков лягушек в водоемах.
Структура населения
На Западно-Сибирской равнине
по результатам классификации сообществ земноводных четко прослеживается три
центра с максимальной плотностью населения (рис.1).
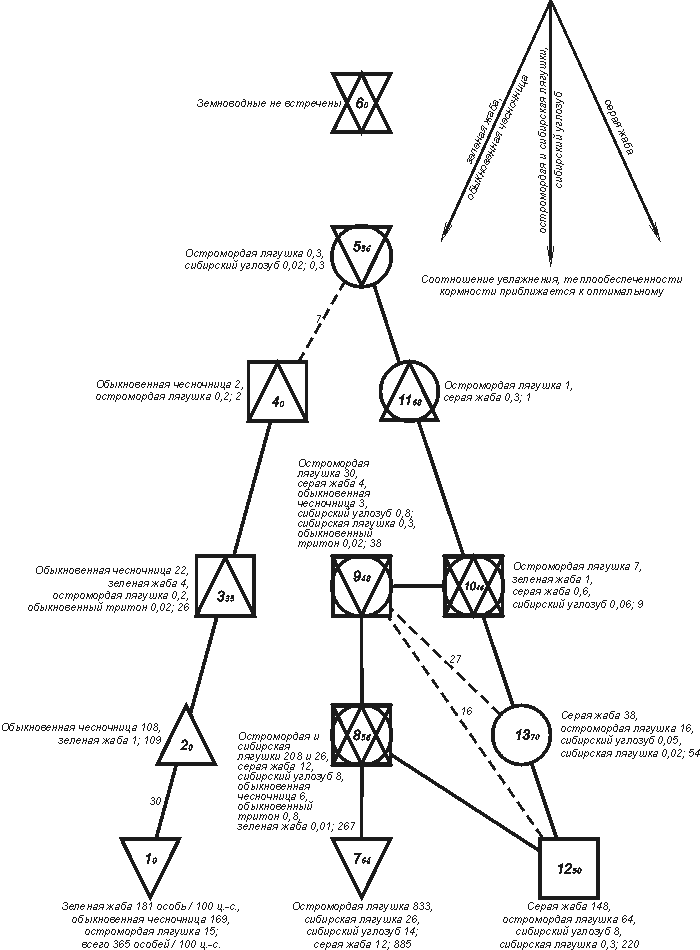
Рис. 1. Пространственно-типологическая
структура населения земноводных Западно-Сибирской равнины.
Схема построена в примерном обратном
масштабе: чем выше межклассовое сходство, тем ближе относительно друг друга
расположены значки с номерами классов и индексами, обозначающими
внутриклассовое сходство. Сила межклассового сходства показана лишь между
первым и вторым классами и в случаях запорогового сходства. Если связь
невозможно отразить в масштабе, она изображена штриховой линией и тоже имеет
оценку сходства.
Квадратами обозначены классы с
преобладанием сообществ лесов нормальной полноты, кружками - тех местообитаний,
где облесенные участки чередуются с
открытыми, а треугольниками - открытых местообитаний (основанием вниз -
обедненных по продуктивности, а вверх - богатых). Рядом со значком класса
приведены встреченные виды и их обилие, а в конце списка - их суммарное обилие.
Варианты населения по классам: 1 - лугов,
займищ и полузаброшенных поселков среди
них в южной части степи; 2 - разнотравно-полынных степей средней и южной
части степной зоны; 3 - сосняков, светлохвойно-мелко-лиственных лесов, посадок,
рямов западной части лесостепи; типчаково-полынных и песчанно-ковыльных степей
средней и южной части степной зоны; 4 - сосняков и полей средней части степной
зоны; 5 - лишайниково-осоково-моховых
тундр и болот южной кустарниковой субарктической тундры; лесотундровых
редколесий и редкостойных лесов долины реки Таз; внепойменных суходольных
местообитаний северной тайги; 6 - территорий, не населенных земноводными
(арктическая, моховая и низкокустарниковая субарктическая тундра; тундры,
бугристые болота и все неприречные светлохвойные леса и редколесья
лесотундровой зоны; песчаные побережья и острова слабосоленого озера Чаны в
лесостепи); 7 - соров и низинных болот
поймы Оби и Иртыша в южной и средней тайге; 8 - низинных болот поймы Оби и
Енисея в северной тайге и подтаежных лесах; лугов, ивняков, осинников поймы Оби
и Иртыша в средней тайге и подтаежных лесах; вне-пойменных низинных открытых
болот южной тайги и подтаежных лесов; подтаежных облесенных низинных болот и
пойменных полей-перелесков; подтаежных заболоченных (хотя бы частично) лесов
долины Тобола; лесо-степных влажных: лесов, лугов, луговых степей, поливных
полей многолетних трав, евтрофных болот, пойм крупных рек; лугов займищ,
сплавин озер и прибрежных тростниковых зарослей в северной и центральной степи;
9 - лесотундровых лесов поймы реки Таз; лугов, ивняков и соров поймы Оби и
Енисея в северной тайге; среднетаежных внепойменных темнохвойных и
мелколиственных лесов и их производных; внепойменных низинных и переходных
болот и пойм притоков крупных рек и Енисея в средней тайге; лугов, ивняков,
осинников поймы Енисея, Оби и Иртыша в южной тайге; внепойменных суходольных
местообитаний южной тайги, подтаежных лесов и лесостепи (кроме лесостепных
поливных полей многолетних трав, а также сосновых и березово-сосновых лесов
западной части подзоны); подтаежных рямов; центральной и северной части степи
(кроме полей, чередующихся с перелесками); 10 - осоковых болот южной
субарктической тундры; лесотундровых комплексных болот и поймы реки Ныда;
северотаежных приречных влажных и полузаболоченных (несветлохвойных) лесов,
редколесий и внепойменных болот; сосняков и светлохвойно-мелколиственных лесов
восточной части лесостепи; сосняков южной части степной зоны; 11 – поймы Оби
(кроме тундр), тампов, приречных редкостойных лесов и редколесий в
лесотундровой зоне; южнотаежных верховых болот; 12 - сосняков их производных и рослых рямов
средней тайги; внепойменных заболоченных облесенных неолиготрофных
местообитаний южной тайги; полей-перелесков центральной и восточной части
северной степи; 13 - среднетаежных верховых
болот; лесостепных березово-осиновых лесов, коллективных садов и
полей-перелесков на месте сосновых приобских боров.
В первом из них (класс 1) численно преобладают
зеленая жаба и обыкновенная чесночница (50 и 44%), во втором абсолютно
доминирует остромордая лягушка (класс 7; 94%); в третьем - серая жаба (класс
12; 87%). От этих центров идет уменьшение обилия перечисленных видов
земноводных в результате снижения теплообеспеченности, продуктивности
биоценозов или увлажнения, а для зеленой жабы и обыкновенной чесночницы - и с
удалением от оптимума ареала и уменьшением к северу степени аридизации. У
обыкновенной чесночницы, кроме того, уменьшение обилия идет и с запада на
восток. У серой жабы и сибирского углозуба оно связано еще и с отрицательным
влиянием половодья на выплод сеголеток, которое тоже снижает обилие лягушек, но
в меньшей степени. Последние имеют в поймах более высокое обилие, чем на
внепойменных суходолах, из-за большей влажности и большего числа пригодных для
выплода водоемов. Тем не менее, на незаливаемых низинных открытых болотах
лягушек больше, чем на заливаемых.
Снижение обилия зеленой жабы
и обыкновенной чесночницы идет в ряду классов 1-6; остромордой и сибирской
лягушек, а также сибирского углозуба - в классах 7-11 и 5; серой жабы - в
классах 12, 13, 10, 11, 5, 6 (состав вариантов по местообитаниям см. в описании
рис. 1). Все ряды сходятся к 6-ому классу, составленному пробами, где
земноводные не встречены. Это все местообитания тундровой и лесотундровой зон,
кроме субарктических лугов, болот, тундр с участием осок, а также лесотундровых
приречных лесов и редколесий. Не встречены земноводные также на песчаных
побережьях и островах слабосоленого озера Чаны. Кроме того, есть еще один
уникальный класс населения земноводных, не представленный на структурной схеме.
Это водные сообщества верховий Оби и Обского водохранилища в пределах
лесостепи. Здесь встречается лишь озерная лягушка. К сожалению учетов ее
численности в водоемах не проведено.
Как видно по составу классов
весьма различные факторы приводят к формированию сходного населения. Так же,
как в большинстве зон и подзон структура населения из-за этого линейна и
совпадает с уменьшением обилия доминирующих видов, правда по равнине в целом
она представлена тремя ветвями, сходящимися к вариантам, где земноводные не
встречены. Таким образом, причины уменьшения могут быть различными, но
приводить к одному и тому же результату - снижению обилия доминантов и соответственно
плотности населения.
Заключение
Таким образом, на
Западно-Сибирской равнине обитает восемь видов земноводных, в том числе три
вида лягушек (остромордая, сибирская, озерная), два вида жаб (серая и зеленая),
сибирский углозуб, обыкновенный тритон и обыкновенная чесночница. Последний вид
встречается преимущественно в западной части подтаежных лесов, лесостепи и степи,
зеленая жаба - только на западе степной зоны и под Новосибирском в лесостепи.
Озерная лягушка нередка на Оби и в Обском водохранилище (лесостепь). Остальные
виды встречаются повсеместно, кроме арктических, северных и средних
субарктических тундр и большей части водораздельных пространств южных
субарктических тундр и лесотундр, где земноводные нами не встречены. Таким
образом, видовое богатство амфибий убывает к северу и югу от лесостепи.
Эмерджентность и целостность
населения земноводных невелика из-за невысокого видового разнообразия.
Пространственно-типологическую структуру их населения определяет, в основном,
степень оптимальности сочетания теплообеспеченности, кормности и увлажнения для
четырех доминирующих видов - остромордой лягушки, обыкновенной чесночницы,
серой и зеленой жаб. Кроме того, прослеживается как положительное, так и
отрицательное прямое и косвенное влияние половодья.
Наиболее благоприятны для
остромордой и сибирской лягушек соры и низинные открытые болота низкой поймы
Оби в южной и средней тайге. Максимальное обилие серой жабы отмечено в сосняках
и сосновых рямах средней тайги, а зеленой жабы и обыкновенной чесночницы - во
влажных лугах и займищах долины реки Тобол в южной степи. Как в пространстве,
так и во времени сравнительно четко прослеживается отрицательная связь обилия
серой жабы и остромордой лягушки. Не исключено, что между ними, кроме некоторых
отличий в предпочтении местообитаний с разной степенью увлажнения, облесенности
и глубины водоемов выплода имеются и конкурентные отношения. В целом, население
земноводных формируется по принципам
обеднения и замещения - независимого и, видимо, зависимого [24].
Обыкновенную чесночницу и
зеленую жабу можно считать относительно термофильными видами, а остальных
мезотермными. В результате влияния внешней cреды (степени оптимальности
гидротермического режима для мезотермов) прослеживается уменьшение плотности
населения амфибий к югу и к северу от подтаежных лесов. Однако хорологические
особенности территории и, видимо, конкурентные отношения серой жабы и
остромордой лягушки приводят к локальному увеличению суммарного обилия
земноводных в средней тайге. Увеличение плотности прослеживается и в степной
зоне за счет проникновения термофилов в западную часть равнины. Расселение их к
северу и к востоку в подтаежных лесах, лесостепи и степной зоне приведет к
почти неуклонному убыванию плотности населения к северу, с одним
отклонением - в средней тайге.
Исследование поддержано по
интеграционным проектом СО РАН № 49 - Биологическое разнообразие Западно-Сибирской
равнины: пространственно-структурная организация, современное состояние и
основные тренды.
Литература
1. Ю.С. Равкин
Пространственная организация населения птиц лесной зоны. Новосибирск: Наука,
1984.
2. В.А. Трофимов, Ю.С.
Равкин Количественные методы в экологии животных. - Л., 1980. 113-115.
3. Ю.С. Равкин, Л.Г.
Вартапетов, В.А. Юдкин и др. Территориальная неоднородность населения
земноводных Западно-Сибирской равнины. - Сибирский экологический журнал, 1995.
- Т.2, № 2. 110-124.
4. О.В. Бурский, А.А.
Вахрушев, С.М. Цыбулин, Фауна и систематика позвоночных Сибири. - Новосибирск:
Наука, 1977а.. 293-300.
5. О.В. Бурский, И.Ю.
Бурская, А.А. Вахрушев, С.М. Цыбулин, Вопро-сы герпетологии. - Л.: Наука,
1977б. 50-51.
6. Ю.С. Равкин, И.В.
Лукьянова, Зоол. журн., 1972. -Т. 61. 929-932.
7. Ю.С. Равкин, И.В.
Лукьянова, Вопросы герпетологии. - Л.: Наука, 1973. 153-155.
8. Ю.С. Равкин, И.В.
Лукьянова, География позвоночных южной тайги Западной Сибири. - Новосибирск:
Наука, 1976.
9. Ю.С. Равкин, Экология.
1976. - № 5. 53-61.
10. Л.Г. Вартапетов, Ю.С.
Равкин, Вопросы герпетологии. - Л.: Наука, 1977. 51-52.
11. Л.Г. Вартапетов, Б.Н.
Фомин, Вопросы герпетологии. - Л.: Наука, 1981. 27.
12. Т.К. Блинова, Вид и его
продуктивность в ареале. - Свердловск. 1984. - Ч.5. 5-6.
13. И.В. Покровская, Вопросы
герпетологии. - Л.: Наука, 1981. 110-111.
14. Л.Г. Вартапетов, И.Г.
Богомякова, И.Г. Маркова, Вопросы герпе-тологии. - Л.: Наука, 1985. 43-44.
15. С.М. Цыбулин, Вопросы
герпетологии. - Киев: Наукова Думка, 1985. 228-229.
16. В.П. Стариков, И.П.
Федосеева, Вопросы герпетологии. - Киев: Наукова Думка, 1989. 242-243.
17. Yu. S. Ravkin, L.G. Vartapetov, V.A. Yudkin et all, Amphibian
Popu-lations in the Commonwealth of Independent States: Current Status and
Declines. - Moskow: Pensoft, 1995. 74-87.
18. Yu.S. Ravkin, V.V. Panov, L.G. Vartapetov et all, Advances in Amphi-bian Research in the Former Soviet Union. - Sofia-Moskow: Pensoft,
1996. - vol. 1. 67-90.
19 Ю.С. Равкин, В.В. Панов,
Л.Г. Вартапетов и др, Вопросы экологии и
охраны позвоночных животных. - Киев-Львов, 1998. - Вып. 2. 49-77.
20. А.Г. Банников, И.С.
Даревский, В.Г. Ищенко и др. Определитель земноводных и пресмыкающихся фауны
СССР. - М.: Просвещение, 1977.
21. Г.С. Золотаренко,
Вопросы герпетологии. - Л.: Наука, 1985. 80-81.
22. В.А. Яковлев, Н.П.
Малков, Вопросы герпетологии. - Киев: Наукова
Думка, 1985. 244-245.
23. М.М. Пикулик,
Земноводные Белоруссии. - Минск: Наука и
техни-ка, 1985.
24. Ю.С. Равкин, Систематика
животных, практическая экология и ландшафтная зоогеография (чтения памяти А.П.
Кузякина). М.: Наука, 1991. 47-48.