УДК: 591.9
Земноводные и пресмыкающиеся равнинных и
предгорно-низкогорных ландшафтов Верхнего Приобья
О.Б. БОРИСОВИЧ, С.М. ЦЫБУЛИН, К.В.
ТОРОПОВ, Б.Н. ФОМИН,
К.В. ГРАЖДАН, И.Н. БОГОМОЛОВА
Институт систематики и экологии
животных СО РАН
630091, Новосибирск, ул. Фрунзе, 11
Сосновые боры
занимают долины Оби и ее притоков в пределах северной и северо-восточной
предгорно-низкогорной части Алтая и проникают в Приобье
на равнину вплоть до подтаежной подзоны в виде почти
непрерывной ленты. Они приурочены к песчаным отложениям и гривисто-лощинному
и дюнному рельефу, благодаря чему, так же как их производные, существенно
отличаются от типичных остепненных и таежных лесов
[1]. В отличие от равнинной лесостепи междуречий и предгорий Алтая, климат
долинных участков более влажный [2]. Кроме того, участие элементов таежной
флоры больше, а неморальных и лугово-степных меньше [3]. Все это определяет их
своеобразие и уникальность. Сосновые боры издавна освоены и коренные формации
на занятой ими территории чередуются с производными смешанными и
мелколиственными лесами, полями и залежами. Поэтому нами рассмотрены
доминирующие по площади местообитания долины Оби как коренные, так и
трансформированные, а также прилежащие к ним ландшафты.
В Верхнем Приобье в настоящее время сравнительно полно исследовано
население птиц, мелких млекопитающих и в меньшей степени земноводных и
пресмыкающихся [4-9]. Поэтому одна из задач наших исследований заключалась в
проведении инвентаризации сообществ земноводных и мелких млекопитающих на тех
ключевых участках разреза по сосново-боровым и прилежащих к ним ландшафтам
Верхнего Приобья, которые были обследованы только по
населению птиц. Та же задача ставилась по пресмыкающимся, учеты которых на этой
территории ранее проводились только в Северо-Восточном Алтае. Общая цель работы
заключалась в выявлении специфики пространственных изменений населения
указанных групп животных при переходе от низкогорных, преимущественно сосновых
лесов и их производных к предгорно-низкогорным ландшафтам долины Бии и далее к
приобским лесостепным сообществам, а также в определении факторов среды,
влияющих на эти изменения.
Материалы и методика
Количественные
учеты земноводных и пресмыкающихся проведены на Северо-Восточном Алтае в
окрестностях предгорных поселков Нижняя Ненинка и Сайдып (Алтайский край) и низкогорных поселков Кебезень и Суучаак (Республика
Алтай), а также в Приобской лесостепной провинции [10] в окрестностях поселков Акутиха и Рассказиха (Алтайский
край, южная лесостепь), Кучук (Алтайский край), Усть-Алеус, Ордынское и Нижнекаменка
(Новосибирская область, северная лесостепь).
Для анализа
использованы материалы, собранные по земноводным в 1976-78, 1989-90, 1993, 1998
и 2000 гг., как правило, за весь период с 16 июля по 31 августа. Обычно учеты
проводились в течение одного лета, только на Нижнекаменском
и Акутихинском участках – по два года. Амфибий
отлавливали 50-метровыми канавками или заборчиками с пятью цилиндрами, на одну
четверть залитыми четырехпроцентным раствором формалина. Для примерного
пересчета показателей обилия на 1 км2 число особей, полученное на
100 цилиндро-суток, умножалось на 300 [11]. Деление
на возрастные группы для остромордой лягушки и серой жабы принято по С.С.
Шварцу и В.Г. Ищенко [12], а для озерной лягушки – по В.И. Гаранину [13] и М.М.
Пикулику [14].
По пресмыкающимся
материалы собраны в мае 1998 и 2000 гг. При прохождении маршрута фиксировалось
расстояние от линии хода учетчика до каждой встреченной особи в момент
обнаружения. Поскольку большинство животных отмечено на полосе 0.5 + 0.5 м (1
м), расчет показателей обилия проводился по выявленным на этой полосе особям.
Если в каком-то местообитании все рептилии встречены вне учетной полосы, за
ширину трансекта принято расстояние до наиболее
удаленного животного. В каждом из обследованных ландшафтных урочищ с учетом
пройдено не менее 5 км. Для сравнимости с данными, полученными по амфибиям,
обилие рептилий увеличено вдвое, т.к. по учетам во второй половине лета в
среднем значения вдвое выше, чем в конце мая - начале июня.
Всего обследовано
64 пространственных варианта населения амфибий и 61 вариант населения рептилий.
Для увеличения надежности показателей обилия и нивелировки влияния межгодовых
колебаний численности подсчитывались средние значения по типологически близким
местообитаниям каждой подзоны и пояса (подпояса), включая результаты двухгодичных учетов. Расчеты
средних проведены без учета соотношения площадей различных ландшафтных урочищ.
Варианты сообществ, для которых отсутствуют данные по обилию одного из этих
классов животных, не вошли в расчеты за исключением поселков, где численность
земноводных и пресмыкающихся крайне низка. При составлении классификации
населения амфибий и рептилий и выявлении его пространственной структуры нами
взяты варианты населения низкогорных поселков, где отсутствуют сведения о
численности пресмыкающихся, и поселков южной лесостепи, по которым нет
материалов по земноводным. При оценке видового состава и плотности сообществ
эти данные в расчеты не вошли. Классификационный и структурный анализ выполнены
по 40 усредненным вариантам населения. Оценка силы и общности связи факторов
среды с неоднородностью сообществ земноводных и пресмыкающихся осуществлена по
исходным неусредненным данным. Классификация
населения проведена с помощью одного из методов кластерного анализа [15] с
использованием коэффициента сходства Жаккара [16] для
количественных признаков [17].
Сведения о
плотности населения земноводных и пресмыкающихся предгорно-низкогорной части и
обилию рептилий в северной и южной лесостепи Верхнего Приобья
опубликованы ранее [9, 18].
Распределение видов
На исследуемой
территории нами встречено три вида амфибий (серая жаба, остромордая и озерная
лягушки) и пять видов рептилий (прыткая и живородящая ящерицы, обыкновенный уж,
обыкновенная гадюка и обыкновенный щитомордник). Последний обнаружен в
сосново-боровом ландшафте низкогорий, где найдена одна мертвая сеголетка. Кроме
того, изучаемые участки равнины входят в границы ареалов сибирского углозуба и сибирской лягушки, а в горах Алтая – узорчатого
полоза [19]. Возможно, отсутствие в учетах первых двух видов связано с
локальностью их распределения в лесостепи Западно-Сибирской равнины.
Серой жабы больше всего в
предгорьях (в среднем 3 тыс. особей/км2). В низкогорьях ее почти
вдвое, в южной лесостепи – вшестеро, а в северной в 73 раза меньше (табл. 1 и
[9, 18]). В ландшафтах северной лесостепи обилие этой жабы выше всего в
сосново-боровом надпойменном и лесополевом (до 0.07
тыс.).
Таблица 1
Земноводные
и пресмыкающиеся равнинной части Верхнего Приобья,
1976-78,
1989-90, 1993 и 2000 гг., особей/км2
|
Подзона,
ландшафт, ландшафтное урочище |
Всего
земноводных и пресмыкающихся |
В том числе*
|
||
Серая жаба
|
Озерная
лягушка |
Остромордая
лягушка |
||
|
Северная лесостепь |
12856 |
35 |
0 |
11770 |
|
Сосново-боровой надпойменный ландшафт |
13912 |
75 |
0 |
10608 |
Березово-сосновые
леса
|
5568 |
0 |
0 |
4060 |
Осиново-березовые леса
|
8877 |
190 |
0 |
3440 |
|
Вырубки |
6355 |
75 |
0 |
1650 |
|
Поля, перелески |
41386 |
0 |
0 |
40140 |
|
Лесополевой надпойменный ландшафт |
4352 |
43 |
0 |
4067 |
|
Поля зерновых и
многолетних трав (поливные и неполивные), луга, перелески |
936 |
50 |
0 |
595 |
|
Кустарники по склонам
долины |
24900 |
0 |
0 |
24900 |
|
Лесолуговой пойменный
ландшафт |
33583 |
0 |
0 |
33500 |
|
Тополево-сосновые леса |
35600 |
0 |
0 |
35400 |
|
Прирусловые ивняки |
52634 |
0 |
+ |
52500 |
|
Луга-выпасы и выгоны |
14400 |
0 |
0 |
14400 |
|
Селитебный ландшафт |
1102 |
0 |
0 |
924 |
|
Поселки |
1180 |
0 |
0 |
1158 |
|
Садово-огородные участки
застроенные |
800 |
0 |
0 |
0 |
|
Южная лесостепь |
40823 |
452 |
3384 |
35011 |
|
Сосново-боровой
надпойменный ландшафт |
13063 |
24 |
474 |
10626 |
|
Сосновые леса |
14597 |
0 |
0 |
13560 |
|
Сосново-березовые леса |
19119 |
0 |
270 |
17370 |
|
Осиново-березовые леса |
16504 |
0 |
270 |
13950 |
|
Залежи |
7333 |
120 |
1680 |
2460 |
|
Луга-покосы |
7652 |
0 |
150 |
5790 |
|
Лесополевой надпойменный ландшафт
(поля зерновых неполивные) |
0 |
0 |
0 |
0 |
|
Болота надпойменные облесенные |
97012 |
1248 |
0 |
94596 |
|
Лесолуговой пойменный
ландшафт |
45978 |
269 |
7002 |
34924 |
Осиново-березовые леса
|
24366 |
540 |
3900 |
11880 |
|
Прирусловые ивняки |
23821 |
60 |
14535 |
6585 |
|
Луга-ивняки |
26934 |
1110 |
150 |
19740 |
|
Луга-покосы |
18520 |
120 |
15900 |
780 |
|
Болота пойменные низинные облесенные и открытые |
51298 |
2340 |
4440 |
43590 |
* сведения по пресмыкающимся опубликованы ранее [18].
+ означает встречу вне учета.
Не встречена она в пойме Оби и в поселках.
Отсутствие этой жабы в пойменных биотопах, по-видимому, связано с перевыпасом скота, при котором часто происходит не только
уменьшение кормности местообитаний из-за вытаптывания и деградации растительности, но и
непосредственное уничтожение этих медлительных животных. В южной лесостепи
серая жаба предпочитает низинные болота (2 и 1 тыс.). Считается, что она
избегает широких пойм и открытых местообитаний, предпочитая лесные массивы [8,
19]. Тем не менее, здесь в пойменных биотопах ее обилие намного выше, чем в
сосново-боровом ландшафте (0.3 и 0.02 тыс.). Исследуемый участок поймы очень
редко затапливается в половодье, при этом имеет лучшую влагообеспеченность и кормность, чем надпойменные биотопы. В предгорьях серой
жабы больше всего на лесостепной равнине (5 тыс.). На заболоченных участках ее
в 8, на мелколиственно-лесной равнине почти в 16 раз меньше. В низкогорьях максимальное
обилие отмечено в сосняках и их производных (2 тыс.), в 3-5 раз ниже оно в переувлажненных местообитаниях и на мелколиственно-лесной
равнине. Не встречена эта жаба в осиново-пихтовой черни.
По местообитаниям северной
лесостепи наиболее часто серая жаба отмечается в надпойменных осиново-березовых
лесах (0.2 тыс.), вдвое реже на вырубках и почти вчетверо в полях с березовыми
перелесками лесополевого ландшафта. В остальных
местообитаниях она не встречена. В южной лесостепи больше всего серой жабы на
пойменных низинных болотах (2 тыс.). На надпойменных болотах и в лугах с
ивняками ее вдвое, в пойменных осиново-березовых лесах вчетверо меньше. Крайне
мало на залежах, в пойменных лугах-покосах и ивняках (0.06-0.1 тыс.). Не
встречена эта жаба в надпойменных лесах и лугах-покосах среди них, а также в
полях. В предгорьях ее обилие убывает от лугов с ивняками и перелесками (7 и 6
тыс.) через поля (0.9), где встречены только расселяющиеся сеголетки, к
березово-осиновым лесам и облесенным низинным болотам
(0.6). В низкогорьях больше всего серой жабы в сосново-березовых разреженных
лесах и в ивняках (7 и 6 тыс.). Впятеро меньше ее в березово-осиновых лесах, в
11-15 раз на облесенных низинных болотах и в березово-осиновых
лесах по гарям и совсем мало на залежах и в сосново-березовых лесах нормальной
полноты.
Таким образом,
серой жабы больше всего в предгорьях и низкогорьях. Наибольшие показатели ее
обилия свойственны мелколиственным лесам в северной лесостепи, низинным болотам
в южной, а также мозаичным по облесенности местообитаниям в предгорьях и
низкогорьях. В целом по Западной Сибири обыкновенная жаба предпочитает в
средней тайге темнохвойно-мелколиственные леса и сосняки, в южной тайге и
подтаежных лесах – облесенные низинные болота [8].
Следовательно, у этой жабы в зависимости от теплообеспеченности, увлажнения и кормности наблюдается смена стаций: в более прогреваемой и
сухой южной лесостепи она чаще встречается на переувлажненной
и пойменной территории, к северу и с увеличением абсолютных высот тяготеет к менее
влажным хвойным и смешанным лесам и мозаичным по облесенности биотопам.
Озерная лягушка – случайно интродуцированный вид. Она попала в водоемы г. Горно-Алтайска
(Республика Алтай) при завозе в качестве лабораторного животного [20] и при
заселении прудов личинками растительноядных рыб, доставляемых из краснодарских
рыбопитомников [21]. Постепенно эта лягушка расселилась от р. Майма вниз по течению, достигнув р. Катунь. Дальнейшее
распространение шло по долине Оби [22]. В ее верхнем течении у поселка Акутиха в 1978 г. отмечены единичные особи этого вида. В
2000 г., судя по отловам в канавки, обилие озерной лягушки возросло здесь в
пойменных биотопах в среднем до 7 тыс. особей/км2.
В северной
лесостепи озерная лягушка встречена вне учета на суше по берегам и на островах
Новосибирского водохранилища. В канавки она попала только в южной лесостепи (в
среднем 3 тыс. особей/км2). Здесь ее больше всего в пойме (в
лесолуговом ландшафте – 7 тыс., на болотном – 4 тыс.) и в 8-14 раз меньше в
надпойменном сосново-боровом ландшафте, где обнаружены только сеголетки и
молодые особи. Из всех обследованных местообитаний больше всего озерная лягушка
тяготеет к прирусловым ивнякам и лугам-покосам (15-16 тыс.). Вчетверо меньше ее
в пойменных осиново-березовых лесах, всемеро – на залежах. Особенно мало
озерной лягушки в надпойменных сосново-березовых и осиново-березовых лесах, на
лугах-покосах, а также в пойме на лугах с ивняками (0.2-0.3 тыс.). Севернее она
отмечена у Томска на городских водоемах, не промерзающих зимой из-за стока
теплых бытовых вод [23].
Таким образом, на
суше озерная лягушка чаще всего попадалась в пойме и лишь иногда во внепойменных облесенных
местообитаниях.
Остромордая
лягушка почти везде многочисленна. Больше всего ее в южной лесостепи (в
среднем 35 тыс. особей/км2). К северу и по мере увеличения
абсолютных высот показатели обилия уменьшаются (в северной лесостепи – 12 тыс.,
в предгорьях – 14 тыс., в низкогорьях – 6 тыс.). В среднем по ландшафтам
северной лесостепи наибольшие значения характерны для пойменных биотопов, втрое
меньшие – для сосново-боровых и в 8 раз – для лесополевых.
Минимальные показатели зарегистрированы в поселках (0.9 тыс.). В южной
лесостепи эта лягушка чаще всего встречается на низинных надпойменных болотах
(95 тыс.), в пойме ее в 2-3 раза, а в сосняках и их производных в 9 раз меньше.
В предгорьях обилие остромордой лягушки убывает в направлении от болот через лесополевые биотопы к мелколиственно-лесной равнине (65; 5
и 2 тыс.). В низкогорьях очень много ее в болотном и мелколиственно-лесном
ландшафте (24 и 19 тыс.) и почти всемеро меньше в сосново-боровых биотопах и в
темнохвойно-мелколиственной черневой тайге.
По местообитаниям северной
лесостепи чаще всего остромордая лягушка встречается в ивняках (53 тыс.). В
полях с перелесками на месте соснового бора, в пойменных тополево-сосновых
лесах и примыкающих к пойме кустарниках по склонам ее насчитывается от 25 до 40
тыс., в пойменных лугах-выпасах – 14 тыс., в надпойменных лесах, на вырубках и
в поселках – от 1 до 4 тыс. особей/км2. Еще меньше ее в полях с
перелесками и не встречена она в садах. В южной лесостепи остромордой лягушки
очень много на низинных болотах (95 тыс. на надпойменных и 44 тыс. на
пойменных). В 4-5 раз меньше ее в пойменных лугах с ивняками и
осиново-березовых, а также в надпойменных лесах, в 12-15 раз – в ивняках, на
залежах и надпойменных покосах. Низкое обилие остромордой лягушки в пойменных
ивняках и на покосах, возможно, связано с вытеснением ее озерной лягушкой,
которой здесь больше, чем где бы то ни было. В предгорьях остромордая лягушка
чаще всего встречается на закустаренных низинных
болотах (65 тыс.). В 9 раз меньше ее в полях, где в канавки ловились только
сеголетки, в 17-20 раз в лугах с перелесками и ивняками, а также в
осиново-березовых лесах и в 145 раз меньше в березово-сосновых лесах. В
низкогорьях она предпочитает облесенные низинные
болота, березово-осиновые леса по гарям и сосново-березовые разреженные леса
(15-24 тыс.) В березово-осиновых и сосново-березовых лесах нормальной полноты,
в осиново-пихтовой черни и ивняках ее обилие в 6-10 раз ниже. Самое низкое оно
в сосняках и на залежах (0.3 тыс.).
Итак, в Верхнем Приобье остромордая лягушка встречена почти повсюду. Больше
всего ее в южной лесостепи. Во всех подзонах она чаще
всего регистрируется на низинных болотах, несколько меньше в пойме и в
надпойменных лесах. Еще меньше в открытых сосново-боровых и лесополевых
местообитаниях и в поселках, что вполне соответствует ранее описанной биотопической приуроченности ее на территории
Западно-Сибирской равнины в целом [8].
Прыткой ящерицы больше всего в
южной лесостепи (в среднем 1 тыс. особей/км2). В северной лесостепи
и низкогорьях ее меньше почти втрое, а в предгорьях в 5 раз. По ландшафтам
северной лесостепи она чаще встречается в сосново-боровом (1 тыс.). В лесополевом и селитебном ее в 6 раз меньше, а в пойме – в
13 раз. В южной лесостепи прыткая ящерица предпочитает пойменные лесолуговые и
в меньшей степени надпойменные сосново-боровые ландшафты (3 и 1 тыс.). В
предгорьях ее обилие максимально на болотах (0.7 тыс.) и вдвое ниже в лесополевом ландшафте. Не встречена она на
мелколиственно-лесной равнине. В низкогорьях эта ящерица обнаружена только в
сосново-боровом ландшафте.
Из всех
местообитаний северной лесостепи прыткая ящерица чаще встречается в
осиново-березовых лесах (2 тыс.). В полях с перелесками среди лесов, в садах и
березово-сосновых лесах ее в 2-3 раза, на вырубках, в полях с березовыми
колками и лесополосами, поливных полях многолетних трав, в пойменных
тополево-сосновых лесах и ивняках в 9-14 раз меньше. Очень мало этой ящерицы в поселках и не
встречена она на выпасах и в кустарниках по склонам. В южной лесостепи
максимальное обилие прыткой ящерицы приходится на пойменные осиново-березовые
леса (8 тыс.), где невысокая сомкнутость крон и, следовательно, хорошая прогреваемость почвы до появления листвы на деревьях
создают весной благоприятные условия для ее обитания. Далее оно снижается в
ряду от залежей, пойменных лугов-ивняков, ивняков и покосов, надпойменных
осиново-березовых лесов (1-3 тыс.), лугов-покосов, сосновых и сосново-березовых
лесов (0.5-0.8 тыс.) к низинным болотам (0.2-0.3 тыс.). Не найдена она среди
обширных полей. В предгорьях на закустаренных
низинных болотах отмечено около 0.7 тыс. особей/км2. Почти вдвое
меньше ее в мозаичных местообитаниях (луга с ивняками и перелесками) и не
обнаружена эта ящерица весной в лесах и полях (одна особь поймана в канавку во
второй половине лета). В низкогорьях прыткая ящерица встречена только в
сосново-березовых лесах. Здесь ее обилие очень велико (5 тыс.).
Таким образом,
прыткая ящерица в равнинной лесостепи предпочитает мелколиственные леса (в
северной – надпойменные, в южной – пойменные) и залежи, в предгорьях – низинные
болота, а в низкогорьях – смешанные разреженные леса. В целом по ареалу она чаще
отмечается в сухих, хорошо прогреваемых местообитаниях, на опушках и обочинах
дорог [19].
Живородящей
ящерицы больше всего в предгорьях и низкогорьях (в среднем 4-5 тыс. особей/км2).
На равнинной части Приобья ее в 10-14 раз меньше.
Аналогичную тенденцию ранее отмечали в украинской части Карпат Н.И. Щербак и
М.И. Щербань [24]. В ландшафтах северной лесостепи эта ящерица
обнаружена только в сосновых борах (2 тыс.). В южной ее обилие убывает от
надпойменных болотных через сосняки и их производные и лесолуговую пойму к
пойменным болотным (0.7; по 0.5 и 0.2 тыс.). В предгорьях живородящая ящерица
предпочитает мелколиственно-лесную
равнину (12 тыс.), но нередко встречается в лесополевом
ландшафте и на низинных болотах (по 2 тыс.). В низкогорьях ее больше всего в
осиново-пихтовой черневой тайге (9 тыс.), вдвое меньше в мелколиственно-лесном
ландшафте, втрое – в сосново-боровом и минимум приходится на болота (0.8 тыс.).
По местообитаниям наибольшее обилие
живородящей ящерицы в северной лесостепи отмечено на вырубках и в
осиново-березовых лесах (3-4 тыс.). В 7-9 раз меньше оно в березово-сосновых
лесах и полях с перелесками среди них. В остальных биотопах эта ящерица не
отмечена. В южной лесостепи она чаще обитает на лугах с ивняками (2 тыс.),
вдвое реже в осиново-березовых лесах, в 3-5 раз на надпойменных низинных
болотах, покосах и в сосняках. Меньше всего ее в надпойменных сосново-березовых
лесах и в пойменных ивняках, на низинных облесенных
болотах и лугах-покосах (0.1-0.2 тыс.). Максимальное обилие живородящей ящерицы
отмечено в предгорных березово-сосновых лесах (16 тыс.). В березово-осиновых
лесах той же подзоны оно вдвое, в лугах с перелесками
и ивняками и на закустаренных низинных болотах в 6-9
раз, а в полях почти в 20 раз ниже. В низкогорьях больше всего этой ящерицы в
осиново-пихтовой черни (9 тыс.). Далее значения убывают от лесов
сосново-березовых разреженных и нормальной полноты, а также березово-осиновых
по гарям (5-6 тыс.) через сосново-пихтово-березовые
леса, ивняки и луга-залежи (2-4 тыс.) к березово-осиновым лесам и облесенным низинным болотам (0.8 тыс.).
Следовательно,
живородящая ящерица предпочитает предгорные и низкогорные местообитания. В
северной лесостепи она чаще встречается на вырубках, в южной – в пойменных
лугах с ивняками, в предгорьях – в смешанных лесах, в низкогорьях – в
осиново-пихтовой черневой тайге. Второе место по предпочитаемости
в большинстве подзон занимают надпойменные
мелколиственные леса.
Обыкновенного ужа больше всего в
предгорьях (в среднем 0.6 тыс. особей/км2). В южной лесостепи его
меньше вчетверо, в северной – в 10 раз и не встречен в низкогорьях. В
ландшафтах северной лесостепи он вдвое чаще встречается в сосново-боровом
(0.1 тыс.), чем в лесополевом. Не нашли его в
пойменных местообитаниях и в поселках. В южной лесостепи в сосново-боровых и
пойменных биотопах обилие ужа одинаково (по 0.2 тыс.) и не обнаружен на
надпойменных болотах и в лесополевом ландшафте. В
предгорьях ужа видели только на мелколиственно-лесной равнине (2 тыс.).
По местообитаниям на севере
приобской лесостепи максимальное обилие обыкновенного ужа выявлено в
надпойменных лесах (0.2 тыс.) и вдвое ниже оно в полях с перелесками лесополевого ландшафта и на вырубках. В южной лесостепи уж
чаще встречается в надпойменных сосново-березовых лесах и в пойменных лугах с
ивняками (0.5-0.7 тыс.). В 3-4 раза меньше его в надпойменных и пойменных
лугах-покосах, а также в ивняках и на низинных болотах поймы. В предгорьях этот
уж обитает в березово-сосновых (3 тыс.) и березово-осиновых лесах (0.7 тыс.). В
остальных местообитаниях не встречен.
Таким образом,
весной обыкновенный уж предпочитает надпойменные леса. Летом же он обычно
тяготеет к биотопам, расположенным еще ближе к рекам, озерам и ручьям [19].
Обыкновенная
гадюка чаще встречается в южной лесостепи (в среднем 0.08 тыс. особей/км2).
Немного меньше ее в предгорьях и в северной лесостепи (0.05-0.06 тыс.) и
вчетверо – в низкогорьях. По ландшафтам северного участка эта гадюка
встречена только в сосново-боровых (0.2 тыс.). В южной лесостепи она
предпочитает пойменные болота (0.3 тыс.), на надпойменных ее вчетверо, в
пойменном лесолуговом ландшафте в 8 раз меньше, а в лесополевом
обыкновенная гадюка не встречена. В предгорьях она попала в учеты только на
мелколиственно-лесной равнине (0.2 тыс.), а в низкогорьях – в сосняках и их
производных (0.03 тыс.).
По местообитаниям в северной
лесостепи больше всего обыкновенной гадюки на вырубках и в осиново-березовых
лесах (0.3-0.4 тыс.) и всемеро меньше в полях с перелесками, расположенными
среди сосново-борового ландшафта. В южной лесостепи ее обилие выше всего на
пойменных низинных болотах (0.3 тыс.), вдвое ниже в ивняках и почти вчетверо на
надпойменных облесенных низинных болотах. В
предгорьях обыкновенная гадюка встречена только в березово-сосновых (0.4 тыс.),
а в низкогорьях в березово-осиновых лесах
(0.2 тыс.).
Итак, в Верхнем Приобье обыкновенная гадюка придерживается надпойменных
мелколиственных и смешанных лесов на севере и в горных районах, а также пойм и
низинных болот в южной лесостепи. Отсутствие встреч в некоторых биотопах
обусловлено низкой численностью вида, а не избеганием этих местообитаний.
Видовое богатство
Наибольшее
количество видов амфибий и рептилий отмечено в лесостепи (7 видов). На
предгорной лесостепной равнине не обнаружена озерная лягушка, а в низкогорьях
кроме нее еще и обыкновенный уж (6 и 5).
В северной
лесостепи видовое богатство населения земноводных и пресмыкающихся убывает от
сосново-боровых надпойменных ландшафтов через лесополевые
к пойменным (6; 4 и 2). В лесополевых местообитаниях
не встречены живородящая ящерица и обыкновенная гадюка, предпочитающие облесенные, более влажные биотопы, а в пойме кроме них –
серая жаба и обыкновенный уж. Возможно, одной из причин бедности пойменной
фауны на этом участке стала повышенная рекреационная нагрузка в связи с его
относительной сухостью и нехваткой лесов в округе [6], а также перевыпас скота, при котором происходит как
непосредственное уничтожение животных, так и исчезновение естественных укрытий.
В южной лесостепи
наблюдается обратная тенденция: количество видов возрастает от надпойменных
террас (на низинных болотах не встречены обыкновенный уж и озерная лягушка, в
сосново-боровом ландшафте – обыкновенная гадюка) к пойме Оби (7 видов). В полях
– единственном исследованном местообитании лесополевого
ландшафта – не встречено ни одного вида обоих классов. Это объясняется
значительной долей территории, подвергшейся распашке, и связанным с этим
отсутствием естественных убежищ, а также сухостью местообитаний.
В предгорьях
население этих животных в открытых и мозаичных биотопах беднее по видам (на
низинных болотах и в лесополевых ландшафтах не
встречены обыкновенные уж и гадюка), чем в лесных местообитаниях (на
мелколиственно-лесной равнине в учетах не отмечена прыткая ящерица,
предпочитающая менее затененные сухие участки). В низкогорье количество видов
амфибий и рептилий уменьшается от сосняков и их производных через
мелколиственно-лесную равнину и низинные болота к осиново-пихтовой черневой тайге.
В первой группе выделов отсутствует обыкновенный уж, во второй кроме него
прыткая ящерица и обыкновенная гадюка. В осиново-пихтовой черни встречены
только остромордая лягушка и живородящая ящерица.
Таким образом,
видовое богатство сообществ земноводных и пресмыкающихся исследуемых ландшафтов
в целом одинаково на равнине, включая предгорья, и снижается в низкогорьях.
Лесостепь отличается по нашим сборам только наличием интродуцированного
вида – озерной лягушки. Из всех ландшафтов равнинной лесостепи наиболее богаты
видами сосново-боровые надпойменные. Фауна пойменных местообитаний южной
лесостепи определенно богаче, чем северной, что обусловлено интенсивным выпасом
скота на обследованных участках последней из подзон. Лесополевые ландшафты и поселки обеднены видами из-за их
сухости, распашки, а также непосредственного уничтожения животных людьми. Такая
же тенденция характерна и для предгорий: лесные биотопы богаче по числу видов,
чем открытые и мозаичные. В низкогорьях фауна обедняется при смене
лесообразующих пород (от мелколиственных и мелколиственно-светлохвойных к мелколиственно-темнохвойным),
а также на склонах по сравнению с террасами.
Итак, амфибии
приурочены в основном к лесным и заболоченным местообитаниям и пойме. Сухие и
обширные распахиваемые участки лесополевого ландшафта
заселяются ими реже. Рептилии предпочитают занимать лесные биотопы, кроме
осиново-пихтовой черни низкогорий, и низинные болота. В отличие от западной
части лесостепи Западно-Сибирской равнины в состав фауны приобских участков не
входит обыкновенная чесночница и зеленая жаба. Не исключено, что это
обусловлено незавершившимся послеледниковым распространением их к востоку и
северу, хотя зеленая жаба случайно завезена и постепенно расселилась под
Новосибирском [25].
Плотность населения и
преобладающие виды
У амфибий по данным учета
канавками наибольшая плотность населения, так же как и видовое богатство,
отмечена в южной лесостепи (38 тыс. особей/км2). В предгорьях их
примерно вдвое, в северной лесостепи – втрое, а в низкогорьях почти впятеро
меньше. Эта тенденция соответствует убыванию обилия остромордой лягушки, на
долю которой приходится 82-99.7 % общего количества земноводных. Второе место
по численности в южной лесостепи занимает озерная лягушка (9 %). Доля серой
жабы возрастает от 0.3 % в северных участках до 18 % в низкогорьях. Это связано
с увеличением ее численности, а не только с уменьшением обилия остромордой
лягушки.
В северной
лесостепи наибольшее суммарное обилие земноводных отмечено в пойменных
лесолуговых ландшафтах (34 тыс.). Почти втрое ниже оно в сосново-боровых надпойменных
и в 8 раз в лесополевых ландшафтах. В поселках
амфибий насчитывается около 1 тыс. особей/км2. Во всех типах
ландшафтов подзоны по численности лидирует остромордая
лягушка, доля которой составляет 99-100 %. Серой жабы значительно меньше. В
пойме и в поселках она вообще не встречена.
В южной лесостепи
амфибий больше всего на низинных надпойменных болотах (96 тыс.). В пойме их
вдвое, а в надпойменных сосново-боровых ландшафтах почти в 8 раз меньше. Численность
остромордой лягушки намного выше, чем у остальных видов (83-99 %). Озерная
лягушка занимает второе место в суммарной плотности населения, но тяготеет к
пойме (0-17 %). Доля серой жабы в общем невелика и составляет 0.2-1 %, и лишь
на пойменных низинных болотах достигает 5 %.
В предгорьях
максимальное суммарное обилие земноводных также приходится на низинные болота
(66 тыс.). В лесополевом ландшафте оно всемеро, а на
мелколиственно-лесной равнине в 35 раз ниже. В последнем случае невысокие
значения определяются низкой численностью сеголеток: их доля в населении не
превышает 8 %, тогда как в других ландшафтах предгорий колеблется от 71 до 91
%. По численности на болотах и в лесах преобладает остромордая лягушка (84-99
%), а в мозаичных биотопах лесополевого ландшафта,
где луга чередуются с ивовыми зарослями и березовыми колками, ее столько же,
сколько серой жабы.
В низкогорьях
наибольшая плотность населения этого класса животных отмечена на низинных
болотах и в мелколиственно-лесных местообитаниях (25 и 19 тыс.). В сосняках и
их производных она ниже вчетверо, а в осиново-пихтовой черни в 8 раз. Доля
остромордой лягушки составляет, как правило, 98-100 %. Только в сосново-боровых
ландшафтах она меньше (62 %), чем у серой жабы.
Распределение
сеголеток в значительной степени зависит от расположения водоемов выплода и поэтому нередко вуалирует зависимость
распределения земноводных от других факторов среды. Если рассматривать
плотность населения без учета сеголеток, то можно сказать, что в северной
лесостепи она максимальна в пойме Оби (18 тыс. особей/км2). В сосново-боровом
и лесостепном ландшафте она, соответственно, в 12 и 17 раз меньше, а минимальна
– в поселках (0.3 тыс.). В южной лесостепи больше всего молодых и взрослых
особей встречается на низинных надпойменных и пойменных болотах и в прочих
пойменных местообитаниях (12-18 тыс.). В надпойменном сосново-боровом ландшафте
их вшестеро меньше. В предгорных районах наибольшее суммарное обилие по этим
возрастным группам отмечено на низинных болотах (6 тыс.), вдвое меньше оно в лесополевом ландшафте, а на мелколиственно-лесной равнине –
в 3.5 раза. В низкогорье плотность населения молодых и взрослых амфибий
максимальна на низинных болотах (7тыс.), а в остальных местообитаниях она в
2.5-3 раза меньше.
Таким образом,
больше всего земноводных отмечено в южной лесостепи. К северу и по мере
увеличения абсолютных высот местности их становится меньше, что совпадает с
уменьшением теплообеспеченности. Также прослеживается сокращение плотности их
населения от низинных болот через незаболоченные
пойменные и сосново-боровые ландшафты к лесополевым и
селитебным. Это направление изменений, отражая снижение оптимальности биотопов
по увлажнению и антропогенной трансформации ландшафтов, связано в основном с
изменением обилия остромордой лягушки. Без учета сеголеток характер изменения
общей плотности населения амфибий сохраняется.
У рептилий максимальное
суммарное обилие характерно для предгорий (6 тыс. особей/км2). Оно
снижается в низкогорьях в 1.5 раза, в южной лесостепи – втрое, в северной –
вшестеро. Это уменьшение показателей определяется прежде всего изменением
обилия живородящей ящерицы, доля которой составляет на равнине 19-50 %, а в
горных районах – около 85 %. В южной лесостепи Приобья,
имеющей большую теплообеспеченность по сравнению с горной частью и более
северными подзонами, первое место по численности
занимает прыткая ящерица (70 %). Доля обыкновенного ужа и обыкновенной гадюки
невелики: у первого вида она колеблется от 6 % в северной лесостепи до 10 % в
предгорьях (в низкогорьях не встречен), у второго – от 0.5 % в низкогорьях до 5
% на северном участке.
В северной
лесостепи наиболее плотно заселен рептилиями сосново-боровое ландшафт (3 тыс.).
Здесь отмечено максимальное обилие всех четырех видов. Преобладают живородящая
и прыткая ящерицы (57 и 33 %). Обыкновенных ужа и гадюки меньше (6 и 5 %). В лесополевом, пойменном лесолуговом ландшафтах и поселках
плотность населения пресмыкающихся в 13-39 раз меньше. В первом из этих
ландшафтов абсолютно доминирует прыткая ящерица (73 %), а в двух последних она
– единственный встреченный вид. В полях-перелесках наряду с ней встречается еще
и обыкновенный уж.
В южной лесостепи
больше всего пресмыкающихся в пойменных лесолуговых ландшафтах (4 тыс.), вдвое
меньше – в сосново-боровых, втрое – на надпойменных болотах и вчетверо – на
пойменных. В первых двух группах биотопов массивы леса с невысокой сомкнутостью
крон и ивняки чередуются с лучше прогреваемыми лугами и залежами. Здесь чаще
остальных видов встречается прыткая ящерица (63-80 %). Второе место занимает живородящая
ящерица, на третьем – обыкновенный уж (13-27 и 5-10 %). Доля обыкновенной
гадюки в пойменном лесолуговом ландшафте не превышает 1 %, а в сосново-боровом
ее не встречали. В местообитаниях со значительным увлажнением лидерами
становятся виды, избегающие сухие открытые пространства: на надпойменной
террасе это живородящая ящерица, в пойме – обыкновенная гадюка (63 и 37 %).
Доля остальных трех видов здесь составляет 0-32 %. По мнению К.В. Граждана с соавторами [9], на болотах, сильно обводненных в
весенний период, пресмыкающихся меньше, чем летом, когда они обсыхают. Во второй
половине лета в этих биотопах они попадаются в канавки значительно чаще, чем в
сухих. Это предположение подтверждается и материалами из южной лесостепи. Кроме
того, у змей наблюдаются сезонные перемещения от мест зимовок, расположенных на
повышенных, хорошо прогреваемых весной участках лесов или склонов, к более
влажным и кормным местообитаниям. Так, П.В. Терентьев и А.С. Чернов [26]
указывают, что ранней весной обыкновенные ужи чаще встречаются в светлых сухих
лесах, летом – в более влажных местах (поймах рек и болотах). Обыкновенная
гадюка, по сведениям З.В. Беловой [27], во время таких же сезонных миграций
перемещается на расстояние до 700 м. Видимо, вскоре после снеготаяния влажность
повсюду вполне достаточна, а межкочечные пространства
на заболоченных территориях залиты водой. Во второй половине лета воды на
болотах уже, как правило, нет, но влажность и кормность
в них существенно выше, чем на высохших суходолах. Это и определяет большую
концентрацию на болотах влаголюбивых видов.
В предгорьях
плотность населения пресмыкающихся максимальна на мелколиственно-лесной равнине
(14 тыс.). Впятеро ниже она на низинных болотах и почти всемеро в лесополевых биотопах. Чаще всего на предгорной равнине
встречается живородящая ящерица (71-85 %). Прыткой ящерицы, предпочитающей на
большей части ареала открытые, хорошо прогреваемые пространства, в предгорьях,
напротив, больше на низинных болотах, чем в лесополевом
ландшафте с лугами, куртинами берез и ив (28 и 16 %), и не встречена она на
мелколиственно-лесной равнине. Обыкновенные уж и гадюка обнаружены только в
последнем из ландшафтов (13 и 1 %).
В низкогорьях
больше всего рептилий в осиново-пихтовой черневой тайге (9 тыс.), в 2-2.5 раза
меньше их в мелколиственно-лесном и сосново-боровом ландшафтах. На низинных
болотах плотность населения пресмыкающихся самая низкая (0.8 тыс.). При этом во
всех группах ландшафтов живородящая ящерица – единственный встреченный вид. Только
в сосново-березовых разреженных лесах к ней присоединяются прыткая ящерица и
обыкновенная гадюка (23 и 1 %).
Итак, суммарное
обилие пресмыкающихся максимально в предгорных районах, убывая к северу и с
увеличением абсолютной высоты местности. Это в большей степени связано с
соответствующим сокращением обилия живородящей ящерицы – лидера по численности
во всех подзонах, кроме южной лесостепи. Наибольшая
плотность населения рептилий отмечается в южной лесостепи в лесолуговых
пойменных, в остальных подзонах – в надпойменных
лесных биотопах. Меньше всего их в лесополевых
местообитаниях, поселках и на низинных болотах, хотя последнее, по-видимому,
относится только к весеннему периоду.
Суммарная
плотность населения земноводных и пресмыкающихся убывает от южной лесостепи к
северной (41 и 13 тыс. особей/км2), а также через предгорья к
низкогорьям (23 и 12 тыс.). Во всех природных подзонах
и поясах по численности доминируют амфибии. Их доля по обилию составляет
60-99.8 %. Только на мелколиственно-лесной равнине предгорий и в низкогорной
осиново-пихтовой черни, где сеголеток лягушек и жаб меньше всего, доминируют
рептилии (76-88 %). Суммарное обилие обоих классов животных убывает от низинных
надпойменных болот (25-97 тыс.) через пойменные местообитания (34-46 тыс.) и
сосново-боровые ландшафты равнинной лесостепи, а также мелколиственно-лесные и
мелколиственно-темнохвойные ландшафты Северо-Восточного Алтая (12-24 тыс.) и
далее сокращается через лесополевые биотопы (4-12
тыс.) к поселкам (1 тыс.). Последнее подтверждает сведения В.А. Ушакова и В.Н.
Гаранина [28] о том, что остромордая лягушка, прыткая и живородящая ящерицы,
которым в Приобье принадлежит основная доля в
плотности сообществ, в населенных пунктах не достигают высокой численности.
Таким образом,
суммарное обилие населения земноводных и пресмыкающихся убывает с уменьшением
увлажнения. Это связано, прежде всего, с преобладанием амфибий, плотность
которых в большинстве биотопов в 2-19 раз больше, чем у рептилий. На уровне подзон и поясов прослеживается влияние теплообеспеченности,
имеющей наибольшую величину в южной лесостепи.
Классификация населения
Население
земноводных и пресмыкающихся рассматриваемого региона по сходству составляющих
его территориальных сообществ делится на 4 типа: оптимальных местообитаний с
низкой и высокой численностью озерной лягушки, субоптимальных
и пессимальных местообитаний. Первые три типа
сообществ сравнительно близки по суммарному обилию и видовому богатству (в
третьем отсутствует интродуцированная озерная
лягушка, которая еще не успела полностью занять исследуемую территорию). Сильно
отличается от них население пессимальных
местообитаний. В эту группу вошли сообщества биотопов, подверженных
значительному антропогенному воздействию (застроенность,
распашка), и низкогорных сосняков. Суммарное обилие в таких местообитаниях
примерно в 20 раз меньше, чем в остальных биотопах первых трех групп сообществ.
Первый и третий типы соответственно можно разделить на подтипы и классы. Ниже
приводится полная классификация.
1. Тип населения
оптимальных местообитаний с низкой численностью интродуцированной
озерной лягушки (равнинных ландшафтов и предгорно-низкогорных низинных болот;
остромордая лягушка – 89 %, прыткая и живородящая ящерицы – 4 и 3 %, озерная лягушка
и серая жаба – 2 и 1 %, обыкновенный уж и обыкновенная гадюка – 0.4 и 0.2 %; суммарное
обилие – 28 тыс. особей/км2; общее число встреченных видов – 7)*.
1.1. Подтип
населения болот и пойменных местообитаний (кроме выпасов, выгонов,
мелколиственных лесов, а также покосов и ивняков южной лесостепи; остромордая
лягушка – 93 %, серая жаба, живородящая и прыткая ящерицы – по 2 % и озерная
лягушка – 1 %, обыкновенный уж и обыкновенная гадюка – 0.3 и 0.1 %; 52 тыс.;
7).
Классы населения:
1.1.1 – равнинных внепойменных и предгорных облесенных
низинных болот (остромордая лягушка – 96 %, живородящая ящерица и серая жаба –
2 и 1 %, прыткая ящерица и обыкновенная гадюка – 0.7 и 0.06 %; 73 тыс.; 5),
1.1.2 – пойменных облесенных низинных болот, лугов, чередующихся с ивняками,
березово-сосновых лесов, а также ивняков северной лесостепи (остромордая и
озерная лягушки – 91 и 3 %, серая жаба, прыткая и живородящая ящерицы – по 2 и
1 %, обыкновенный уж и обыкновенная гадюка – 0.5 и 0.1 %; 42 тыс.; 7).
1.2. Подтип
населения внепойменных лесов, зарастающих
вырубок, покосов, залежей и полей, перелесков сосново-борового ландшафта,
кустарников на склонах, пойменных мелколиственных лесов, выпасов, выгонов, а
также низкогорных облесенных низинных болот
(остромордая лягушка – 82 %, прыткая и живородящая ящерицы и озерная лягушка –
8; 5 и 3 %, серая жаба, обыкновенный уж – по 0.7 % и обыкновенная гадюка – 0.3
%; 17 тыс.; 7).
2. Тип населения
оптимальных местообитаний с высокой численностью интродуцированной
озерной лягушки (пойменных покосов и ивняков южной лесостепи; озерная и
остромордая лягушки – 72 и 12 %, прыткая ящерица – 9 %, обыкновенный уж,
живородящая ящерица – 0.8 и 0.6 %, серая жаба и обыкновенная гадюка – по 0.4 %;
21 тыс.; 7).
3. Тип населения субоптимальных местообитаний (предгорно-низкогорных ландшафтов,
кроме болот; остромордая лягушка, живородящая ящерица и серая жаба – 39; 36 и
18 %, прыткая ящерица и обыкновенный уж – 4 и 2 %, обыкновенная гадюка – 0.4 %;
15 тыс.; 6).
Подтипы населения:
3.1 –
предгорно-низкогорных полей и лесов, исключая сосняки (остромордая лягушка и
живородящая ящерица – 43 и 41 %, серая жаба, прыткая ящерица и обыкновенный уж
– 8; 4 и 3 %, обыкновенная гадюка – 0.5 %; 15 тыс.; 6),
3.2 – предгорных и
низкогорных лугов, чередующихся с ивняками и березово-осиновыми колками (серая
жаба, остромордая лягушка и живородящая ящерица – 51; 26 и 21 %, прыткая
ящерица – 3 %; 13 тыс.; 4).
4. Тип населения пессимальных местообитаний (поселков, садов, низкогорных
залежей и сосняков, а также равнинных полей и полей-перелесков в пределах лесополевого ландшафта; остромордая лягушка, живородящая и
прыткая ящерицы – 40; 33 и 20 %, серая жаба и обыкновенный уж – 5 и 1 %; 0.7
тыс.; 5).
Пространственно-типологическая
структура
Под
пространственно-типологической структурой животного населения мы понимаем
основные направления изменений населения животных в пространстве структурообразующих
факторов среды [29]. Графы сходства сообществ земноводных и пресмыкающихся
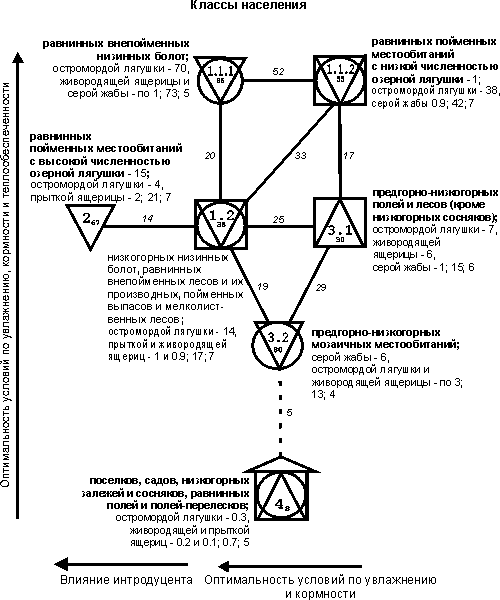
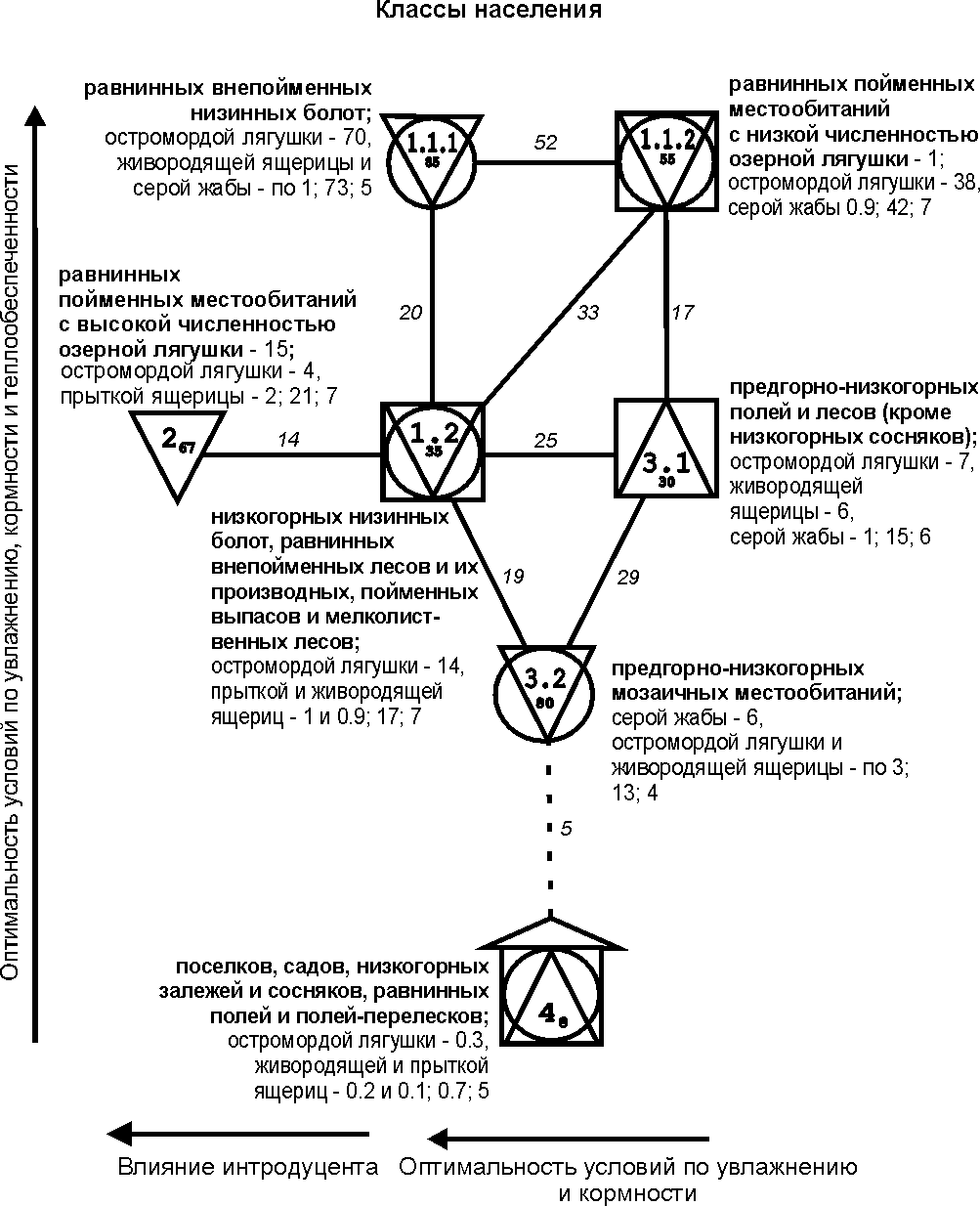
построены при пороге значимости выше 11 единиц (рис. 1-3).
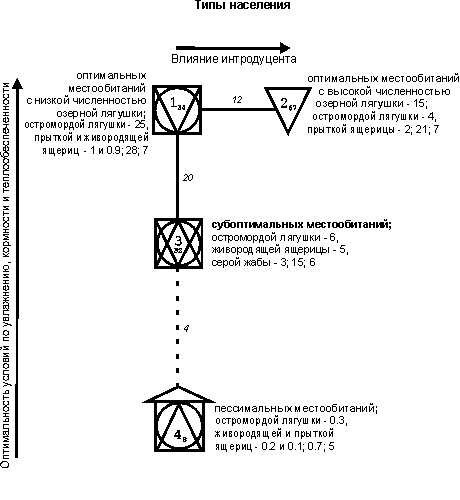
{kind=link}
Рис.
1. Пространственно-типологическая структура
населения земноводных и пресмыкающихся (на уровне типа) в приобских лесостепных
и предгорно-низкогорных ландшафтах.
На рис. 1-3
квадратами обозначены сообщества территорий с преобладанием лесов, кружками –
мозаичных по облесенности участков, треугольниками – открытых местообитаний
(основанием вниз – обедненных по продуктивности, вверх – богатых), «домиком» –
застроенных территорий. Рядом со значком класса приведены три первых по численности
вида и их обилие, в конце списка – суммарное обилие (тыс. особей/км2)
и общее число встреченных видов. Индекс у номера класса обозначает
внутриклассовое сходство, между классами – межклассовое; менее значимые связи
отмечены прерывистой линией.
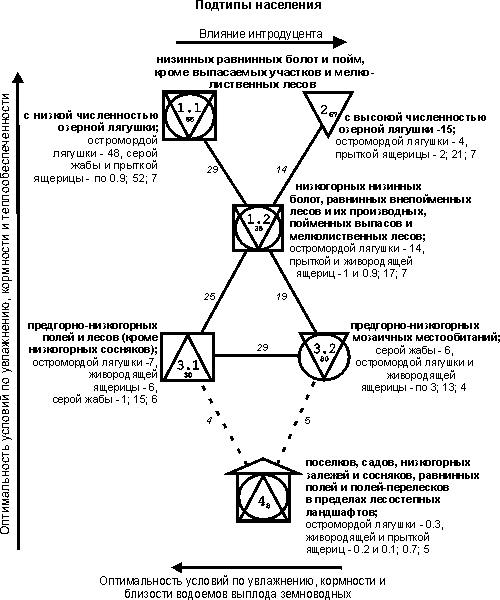
{kind=link}
Рис.
2. Пространственно-типологическая
структура населения земноводных и пресмыкающихся (на уровне подтипа) в
приобских лесостепных и предгорно-низкогорных ландшафтах.
{kind=link}
Рис. 3. Пространственно-типологическая структура населения
земноводных и пресмыкающихся (на уровне класса) в приобских лесостепных и
предгорно-низкогорных ландшафтах.
Структурный граф на уровне типа населения
отражает два направления изменений: от оптимальных биотопов с низкой
численностью озерной лягушки (1-ый тип) через сообщества субоптимальных
местообитаний (3-й) до населения пессимальных
участков (4-й). Отклонение от этого основного ряда представлено типом населения
оптимальных биотопов, но с высоким обилием озерной лягушки (2-й). В первом ряду
прослеживается снижение значений плотности и видового богатства населения от
равнинных заболоченных, пойменных и лесных биотопов через предгорно-низкогорные
ландшафты к лесополевым, распаханным местообитаниям и
застроенным территориям. В последний тип также вошли сообщества низкогорных
сосняков. Своеобразные «засушливые» условия в них создаются вследствие того, что
сосновые боры произрастают на сложенных песчаными отложениями террасах, где
выпадающие осадки быстро стекают или просачиваются [30]. Эдификатором
сходства выступает остромордая лягушка, обилие которой убывает от первого к
четвертому типу. С продвижением от заболоченных и пойменных биотопов через леса
к полям и лугам в пределах лесополевых ландшафтов
прослеживается снижение оптимальности из-за уменьшения увлажнения и сокращения
продуктивности сообществ, и в частности количества доступных для питания беспозвоночных.
С увеличением абсолютных высот местности от равнины к низкогорьям постепенное
снижение численности определяется понижением теплообеспеченности. Отрицательное
воздействие на сообщества земноводных и пресмыкающихся оказывают застройка и
распашка территории. Таким образом, в первом ряду четко прослеживается снижение
плотности и видового богатства населения при ухудшении условий обитания из-за
соответствующего уменьшения увлажнения, кормности и
теплообеспеченности местообитаний, а также увеличения антропогенной нагрузки.
Другой тренд
отражает незавершенность расселения и влияние в южной лесостепи интродуцированного вида – озерной лягушки. Вероятно, этот
вид имеет конкурентное преимущество перед остромордой лягушкой и серой жабой и
снижает их численность. По сведениям С.Л. Кузьмина [31], особи озерной лягушки
активнее кормятся наземными и водными объектами, а также икрой амфибий,
поскольку передвигаются значительно быстрее остромордой лягушки и особенно
серой жабы. Доля головастиков и сеголеток от общего числа жертв этой лягушки
обычно невелика. Только в период метаморфоза крупные особи частично
переключаются на питание молодью амфибий, и они иногда встречаются почти во
всех желудках взрослых. В некоторых водоемах ими выедается до 98-99 % особей,
недавно завершивших метаморфоз. Судя по нашим учетам, присутствие озерной
лягушки вызывает снижение численности других амфибий.
Тренды,
прослеженные на уровне типа, на подтиповой структуре
сохраняются (см. рис. 2). Общее обилие земноводных и пресмыкающихся уменьшается
от равнинных низинных болот и пойм (кроме входящих в подтип 1.2) через
низкогорные низинные болота, равнинные внепойменные
леса, пойменные выпасы и мелколиственные леса, далее через предгорно-низкогорные
местообитания до биотопов с пессимальными условиями,
вошедших в четвертый подтип. В отличие от остальных местообитаний поймы, в
мелколиственных лесах на втором месте по численности стоит прыткая ящерица, а
живородящая не встречена. Обилие серой жабы невысокое. Пойменные выпасы
исследуемого участка характеризуются заметной деградацией травяного покрова
[6]. В этом ряду четче, чем на подтиповой структуре,
прослеживается сокращение плотности и видового разнообразия сообществ по мере
уменьшения оптимальности условий по увлажнению, кормности
и теплообеспеченности. Помимо этого, суммарное обилие и число встреченных видов
снижаются при переходе от естественных биотопов через пойменные выпасы к
поселкам и полям, что определяется ухудшением условий обитания из-за увеличения
антропогенного воздействия.
Кроме того,
проявляется дополнительный тренд. В типе субоптимальных
местообитаний выделяются два подтипа: предгорно-низкогорных полей и лесов
(кроме низкогорных сосняков; 3.1) и мозаичных биотопов, куда вошли сообщества
предгорных лугов, чередующихся с колками и ивняками, и пойменных ивняков (3.2).
Поля предгорий, вошедшие в подтип 3.1, отличаются от приобских тем, что они
невелики по площади и перемежаются с лесами и небольшими западинами с ручьями,
что увеличивает численность амфибий и рептилий. Однако эти поля избегаются
молодыми и взрослыми земноводными, а сеголетки встречаются там только при
расселении. С продвижением от лесов к лугам с ивняками и колками сокращение
суммарного обилия и видового богатства сообществ определяется снижением
оптимальности условий существования по увлажнению и кормности.
Увеличение суммарной численности земноводных и пресмыкающихся от мозаичных
биотопов к полям связано с близостью последних к водоемам выплода.
На уровне класса
(см. рис. 3) население амфибий и рептилий делится на 7 групп. На равнинных внепойменных низинных болотах (класс 1.1.1) плотность
населения максимальна, хотя видовое богатство ниже (нет обыкновенного ужа и
озерной лягушки), чем на пойменных покосах, лугах в сочетании с ивняками,
ивняках и хвойно-лиственных лесах (1.1.2). В этом ряду увеличение суммарного
обилия связано с увеличением оптимальности условий в первую очередь для амфибий
при возрастании увлажнения, которое выше на заболоченных участках, чем в
пойменных биотопах. В остальном представленные на подтиповой
структуре направления изменений населения земноводных и пресмыкающихся
различных местообитаний сохраняются.
Информативность
структурных представлений на уровне типа населения, оцененная по доле учтенной
дисперсии коэффициентов сходства, составляет 23 %. На уровне подтипа и класса –
соответственно, 14 и 11 % дисперсии, а по всем таксонам вместе – 42 % (коэффициент
регрессии – 0.65).
В целом по
лесостепи Западной Сибири выделяются два направления изменений в населении
земноводных. В первом случае сокращение численности идет от влажных не
распахиваемых ежегодно местообитаний через суходольные открытые и мозаичные к сосновым
и сосново-березовым лесам. Этот тренд обусловлен соответствующим уменьшением
увлажнения. Второй ряд демонстрирует провинциальные различия сообществ амфибий Тоболо-Ишимского и Обь-Иртышского
участков, связанные в основном с обитанием на западе обыкновенной чесночницы.
Снижение оптимальности увлажнения и кормности вызывает
сокращение плотности населения во всех подзонах
Западно-Сибирской равнины, а, начиная от южной тайги, к югу сказывается влияние
поемности, от подтаежных лесов – провинциальности, в
степной зоне – ксерофитизации [7].
Итак, при
рассмотрении структуры населения земноводных и пресмыкающихся
предгорно-низкогорных ландшафтов Верхнего Приобья
основные направления его изменений, прослеживающиеся в целом на
Западно-Сибирской равнине и связанные с уменьшением увлажнения, кормности, провинциальности, сохраняются. Кроме них
выделяются дополнительные тренды, зависящие от проникновения интродуцированного вида (озерной лягушки), а также от
изменения теплообеспеченности при увеличении абсолютных высот местности над
уровнем моря.
Организация населения
В целом по
Верхнему Приобью
наиболее существенное по силе и общности влияние на формирование населения
земноводных и пресмыкающихся оказывает антропогенная трансформация ландшафтов:
распашка и застроенность территории (11 и 6 %
учтенной дисперсии, в целом 12 %). Несколько менее значимы увлажненность и кормность местообитаний, а также облесенность
и состав лесообразующих пород (по 9; 6 и 5 %). Существенно меньше сказывается
влияние поемности (3 %), которая влияет на население
амфибий и рептилий двояко. С одной стороны после половодий в пойме появляется
большое количество временных водоемов, удобных для откладки икры земноводных, а
высокая влажность по сравнению с суходолами увеличивает продуктивность
биоценозов. С другой стороны затопление и заиливание прибрежных участков
водоемов выплода сокращает выживаемость икры и
молоди. Отрицательное воздействие половодий особенно сильно сказывается на
численности сибирского углозуба, серой жабы и
пресмыкающихся.
Еще меньшее
влияние на сообщества этих групп животных оказывает зональность и изменение
абсолютных высот местности (по 2 %). Этими факторами среды в целом учитывается
19 % дисперсии, а природными режимами по классификации населения – 30 % и 42 %
по структурам. Всего всеми факторами и режимами учитывается 48 % дисперсии
населения, отраженной коэффициентами сходства (коэффициент регрессии – 0.69).
При изучении
населения земноводных Западно-Сибирской равнины в целом на первое место выходит
зональность, что связано со значительными изменениями теплообеспеченности с
продвижением с севера на юг. Антропогенное влияние, увлажнение и кормность оказываются менее существенными [8]. Остальные
факторы имеют почти такое же проявление силы и общности связи с неоднородностью
сообществ земноводных, как и для населения приобской лесостепи и
предгорно-низкогорных ландшафтов.
Заключение
На территории
приобских и бийских сосново-боровых и прилежащих к
ним ландшафтов нами встречено 3 вида амфибий и 5 видов рептилий. Плотность и
видовое богатство их населения постепенно убывает от внепойменных
низинных болот через пойменные местообитания, внепойменные
приобские леса и предгорно-низкогорные поля, леса и мозаичные биотопы к
поселкам, садовым участкам, низкогорным залежам и соснякам, а также приобским
полям и полям-перелескам лесополевого ландшафта.
Суммарное обилие уменьшается с увеличением абсолютных высот местности и
снижением теплообеспеченности. При удалении от пойм такое снижение объясняется
уменьшением оптимальности местообитаний при соответственном снижении их
увлажнения и кормности. Немаловажное значение в
сокращении плотности населения представленных групп позвоночных приобретает
антропогенное воздействие (застроенность, распашка).
По суммарному обилию среди двух представленных классов позвоночных во всех
группах сообществ лидируют амфибии, особенно остромордая лягушка. Это прежде
всего связано с близостью поймы Оби и притоков Бии, наличием временных и
постоянных водоемов выплода, а также с меньшей
сухостью климата по сравнению с лесостепным междуречьем.
Литература
1. П.Л. Горчаковский, Бот. журн., 1949, 34: 5, 524-538.
2. Г.Н. Огуреева, Ботаническая география Алтая, М., Наука, 1980.
3. Е.И. Лапшина,
Растительный покров Западно-Сибирской равнины, Новосибирск, Наука, Сиб. отд-ние, 1985, 120-125.
4. А.С. Николаев, Биологические ресурсы поймы Оби, Новосибирск, Наука, Сиб. отд-ние, 1972, 60-121.
5. Ю.С. Равкин, Птицы Северо-Восточного Алтая, Новосибирск, Сиб. отд-ние, 1973.
6. Б.Н. Фомин, Проблемы зоогеографии и истории фауны, Новосибирск, Наука, Сиб. отд-ние, 1980, 273-286.
7. Ю.С. Равкин, Л.Г. Вартапетов, В.А. Юдкин и др., Сиб. экол. журн., 1995, 2: 2, 110-124.
8. Ю.С. Равкин, В.В. Панов, Л.Г. Вартапетов и др., Вопросы экологии и охраны позвоночных животных, вып. 2, Киев-Львов, 1998, 49-77.
9. К.В. Граждан, К.В. Торопов, У.Ю. Веряскина, Животный мир Алтае-Саянской горной страны, Горно-Алтайск, 1999, 43 - 50.
10. Западная Сибирь, М., Изд-во Акад. наук СССР, 1963.
11. Ю.С. Равкин, И.В. Лукьянова, География позвоночных южной тайги Западной Сибири, Новосибирск, Наука, Сиб. отд-ние, 1976.
12. С.С. Шварц, В.Г. Ищенко, Пути приспособления наземных позвоночных
животных к условиям существования в субарктике, т. 3, Свердловск, Труды ин-та экологии растений и животных, 1971, вып. 79.
13. В.И. Гаранин, Земноводные и пресмыкающиеся Волжско-Камского края, М., Наука, 1983.
14.
М.М. Пикулик, Земноводные Белоруссии, Минск, Наука и
техника, 1985.
15. В.А. Трофимов, Ю.С. Равкин, Количественные методы в экологии животных, Л., 1980, 113-115.
16.
P. Jaccard, Bull. Soc. Vaund.
Sci. Nat., 1902, 38, 69-130.
17. Р.Л. Наумов, Птицы в очагах клещевого энцефалита Красноярского края, Автореф. дис. … канд. биол. наук, М., 1964.
18. О.Б.
Борисович, С.М. Цыбулин, К.В. Торопов, Вопросы
герпетологии, Пущино-Москва, 2001, 41-43.
19. А.Г. Банников, И.С. Даревский, В.Г. Ищенко, А.К. Рустамов, Н.Н. Щербак, Определитель земноводных и пресмыкающихся фауны СССР, М., Просвещение, 1977.
20. В.А. Яковлев, Н.П. Малков, Вопросы герпетологии, Л., Наука, Ленингр. отд-ние, 1985, 244-245.
21. К. Филиппов, Рыбоводство и рыболовство, 1982, 7, 5.
22. В.А. Яковлев, Животный мир Алтае-Саянской горной страны, Горно-Алтайск, 1999, 175-214.
23. В.Н. Куранова, Вопросы герпетологии, Пущино-Москва,
2001, 147-149.
24. Н.И. Щербак, М.И. Щербань, Земноводные и пресмыкающиеся Украинских Карпат, Киев, Наук. думка, 1980.
25. Г.С. Золотаренко, Вопросы герпетологии, Л., Наука, Ленингр. отд-ние, 1985, 80-81.
26. П.В. Терентьев, А.С. Чернов, Определитель пресмыкающихся и земноводных, М.-Л., Сов. наука, 1949.
27. З.В. Белова,
Вопросы герпетологии, Л., Наука, Ленингр. отд-ние,
1973, 35-36.
28. В.А. Ушаков, В.И. Гаранин, Вопросы герпетологии, Л., Наука, Ленингр. отд-ние, 1973, 185-187.
29. Ю.С. Равкин, Птицы лесной зоны Приобья, Новосибирск, Сиб. отд-ние, 1978.
30. Г.С. Самойлова, Природа очагов клещевого энцефалита на Алтае (северо-восточная часть), Новосибирск, Наука, Сиб. отд-ние, 1967, 5-18.
31. С.Л. Кузьмин,
Земноводные бывшего СССР, М., Товарищество научных изданий КМК, 1999.