SPATIAL-TYPOLOGICAL
HETEROGENEITY OF BIRD COMMUNITY OF EAST-EUROPEAN AND WEST-SIBERIAN PLAINS
E.S. Ravkin*1, L.G. Vartapetov2,
K.V. Toropov2, Yu.S. Ravkin2,
S.M. Tsybulin2, V.S. Zhukov2,
I.V. Pokrovskaja3, I.I. Byshnev4.
S.K. Kochanov5
1. Center of ecologic-geographic developments SIA
“Radon”, Moscow, 127562, Russia, ravkin_e@mtu-net.ru;
2. Laboratory of zoological monitoring Institute of
Animal Systematics and Ecology Siberian Branch of Russian Academy of Sciences,
Novosibirsk, 630091, Russia, zm@eco.nsc.ru;
3. Laboratory of biogeography – IG RAS, Moscow,
109117, Russia, pokrov@biogeo.msk.ru;
4. Berezinsky reserve, Domzheritzy, 211188, Byelorussia;
5. NB Komi NC UrORAS, Syktyvkar, 167610, Russia
*To whom correspondence should be addressed. Russia,
127562 Moscow, Altufievskoye Road 30-229
Key words: East-European and West-Siberian Plains, zoogeography, classification, bird population, ornithocomplexes, bird community, population density, biodiversity, zone, subzone.
Abstract
Spatial-typological structures of bird
population have been determined during the first half of summer in the
East-European plain. The data obtained were compared with that of similar
structure in the West-Siberian plain and between the two plains. It was shown
the close similarity in the revealed structures and their organization.
Introduction
By spatial-typological structure of the
population of bird heterogeneity is meant the general property of territorial
differences of ornithocomplexes, i.e. main trends of changes in bird
communities in space of structure-forming environmental factors, including
animal interrelations.
Materials and methods
See in the previous article E.S. Ravkin et
al.
Results and discussion
Spatial-typological heterogeneity of animal
population may be reflected by hierarchical classification of the communities
and structural graph, i.e. by the classification, reflecting taxa
interrelations on the coefficient of similarity. Such population structure of
birds in the East-European plain on the level of the community type reflects
the presence of three systems (zonal rows). The first of them involves natural
and field communities, the second – towns, settlements, dumps and ruderal
ornithocomplexes, and the last one – water-water-adjacent communities
(fig. 1). The first row belongs to the first revealed system, and is
presented by the trend on the gradient of latitudinal heat supply (zonality)
(type 1, 2, 5, 6). On subject considerations, it is the most important, though
formally the incoming types only in one case are significantly connected (types
5 and 6). Weak threshold-overlapped connection is traced between the boreal and
flood-plain forest-steppe types (2 and 5), while the tundra and boreal types (1
and 2) are slightly similar and connected by the population of high bogs. In
real space and in other population systems the alteration of zones and thus of
their corresponding types of the communities doesn’t cast any doubt, therefore
suggested interpretation is conceptually preferable.
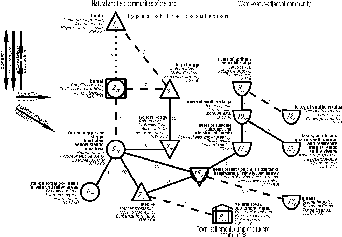
{kind=link}
Fig. 1. Spatial-typological
structure of bird community of East-European plain in the first half of summer
on type level. Schemes 1-4 are approximately reverse scaled: the higher
interclass similarity, which is located between classes, the closer relative to
each other marks with numbers of types and subtypes and indices, representing
the interclass similarities. Inter –and intraclass similarities are estimated
as the mean similarity coefficients of the appropriate community variants. If
similarity lay over threshold or the connection is not possible at scale, it is
shown by dotted and hatching lines and also has the estimation of similarity.
Squares are classes with dominated communities of forests, circles – habitats,
where afforestated areas are alternated with the open ones, triangles ‑
open habitats (base downward ‑ with poor productivity, upward – with rich
productivity), half-circle – reservoirs and rivers, "houses" – towns
and villages, rectangles – heaps. Close to the class mark are three first on
number species, population density and number of the background species.
Thus, the tundra population of birds in the
East-European plain is changed by the boreal one, followed by ornithocomplexes
of the forest steppe and steppe flood-plain willow stands and meadows, then by
steppe extraflood plain forests and felled areas, followed by the steppe-like
ornithocomplexes, which location is dual. On the one hand it is undoubtedly zonal
type, on the other due to the impoverished field population, involved in this
type, it forms deviation from the main row. It causes the graph trend
divarication in the lower part. In zonal row the population density is maximal
in steppe extraflood plain forests and felled areas, diminishing to the north
up to the tundra type. The number of the background species changes almost the
same way as the density, but the highest records are characteristic of the
boreal type population due to the greater territory it occupies. The population
of the high-bog and marsh types also shows the deviation from the main row.
Soil swamping and the impoverished mineral nutrition of phytocenoses decrease
their productivity, thus, lowering the population density and the background
abundance of the population. Changes in the dominant species composition from
the north to the south resulted in the replacing of tundra species by the
forest and margin forest species or by more southern species, while in
deviations and in town, settlement, dump and ruderal and water-water-adjacent
communities the portion of bogy, meadow and water-water-adjacent birds
increases. Similar picture occurs with anthropogenic transformation of original
landscapes, synanthropic plants. The structure graph, based on the level of
subtype, factually confirms in details the records, traced on the type level
(fig. 2). The informativity of subtype records is twice as high (44%
dispersion of similarity coefficient of ornithocomplexes). Here the main zonal
row of changes consists of extraflood-plain tundra and swamp ornithocomplexes
(first of tundra zones and then of the forest tundra). It is followed by the
population of light forests, forests, felled areas and flood- plains of tundra
zone, forest tundra and northern taiga. Then, the forest ornithocomplexes (from
the middle taiga to the forest steppe) are changed by the population of steppe
extraflood-plain forests and felled areas. On similarity they are followed by
the bird communities of flood-plain forest steppe and steppe willow stands in
combination with meadows and, finally, by that of extraflood-plain meadows,
steppes, fields and shrubs of the forest steppe and steppe zones. As this takes
place the weak connection is traced between the communities of the forest
tundra bogs and tundra (subtypes 1.1 and 1.3) and forest ornithocomplexes (2.1,
2.2 and type 6), then between the letter and the communities of willow
stand-meadow-steppe types (5 and 7), which are not divided into subtypes.
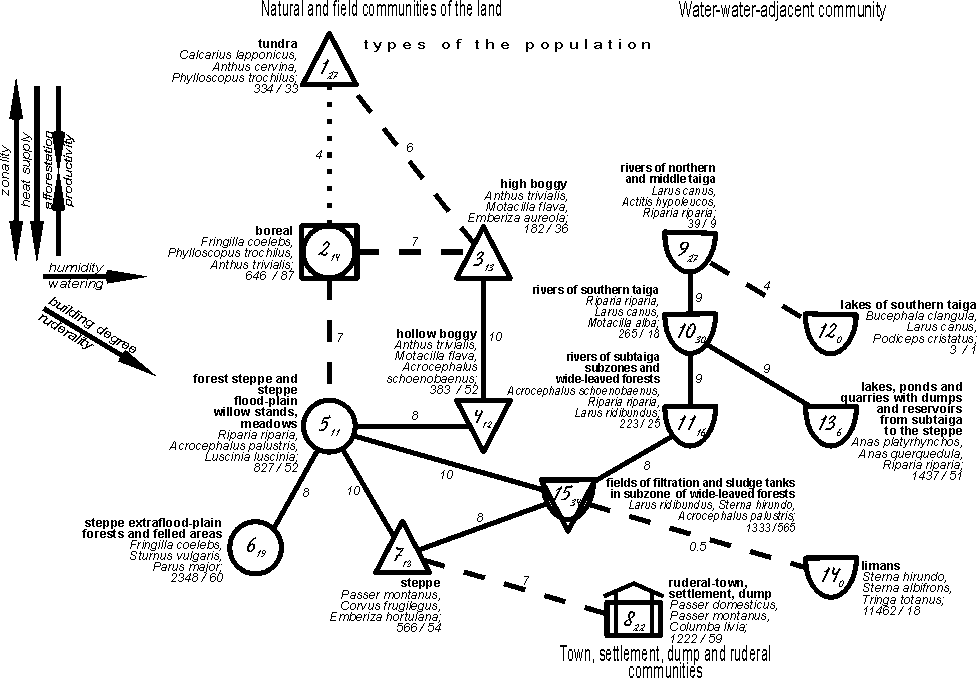
{kind=link}
Fig. 2. Spatial-typological structure
of bird community of East-European plain in the first half of summer on subtype
level.
In zonal row of the population subtypes the
bird abundance almost constantly increases to the south from 343 to 2348
specimens per km2, decreasing in the steppe type up to 566 specimens per km2
with the exception of the population density in light forests, felled areas and
flood-plains of tundra zone and forest tundra, being somewhat smaller of the
predicted. Usually in flat northern taiga and mountain light forests of Siberia
birds are more abundant than in tundra. The european property is associated
with the worming effect of the North-Atlantic current, more influencing the
ocean tundra, than the forest tundra, located at the great distance from the
sea coast. Maximum background abundance is characteristic of the forest subtype
community of this row (57-80 species). They are less abundant in the most
southern open and shrubby communities (52-54) and considerably less number is
observed in ornithocomplexes of tundra type (23-31). The dominant composition
in the row is changed from the tundra species to the forest tundra and then to
the forest, flood plains and species of open forest steppe and steppe habitats,
that is in agreement with zonal alteration of vegetation and is determined by
it.
Coincidentally with the described process,
zonal row of changes takes place in the communities of habitats, considered as
intrazonal. In tundra zone and the forest tundra it is the population of the
extra flood-plain meadows (subtype 1.2), in the forest zone and the forest
steppe – bogs (types 3 and 4) and the forest-meadow-field habitats (subtypes
2.3). In these types and subtypes the total bird abundance is always smaller
than in the habitats of the main zonal row, that is probably caused by the
decreased productivity of phytocenoses. In the forest zone, it is typical for
high, transitional and adjacent mashes, which trophic-capacities are lower,
than in the forest and trophically rich marshes and meadows, especially
flood-plain meadows. In forest-meadow-field habitats, the trophic capacity is
lower due to the development of monoculture agrocenoses and probably an
intensive haying and overgrazing of cattle in meadows. In Siberia the meadows
are maintained better due to not so actively developed agriculture, and the
investigated marshes, especially, flood-plain marshes, are trophically richer
and thus more preferable for birds. Therefore, in the West-Siberian plain the
meadow-bogy variants of the population have high density and deviate from the
main row due to the higher (but not lower as in Europe) productivity (food
reserve). The dominants’ composition in the row of deviations also coincides
with zonal change of vegetation. Meadow-tundra birds are changed by the
bogy-meadow and species of mosaic plants and shrubs. The population density and
the abundance of the background species in this row constantly increase from
the north to the south.
This row is connected with town,
settlement, dump and -ruderal and water-water-associated communities by the
bird community's filtration fields and sludge tanks. Sludge tanks and
filtration fields due to their high trophic capacity are very attractive for
many birds of meadow-bogy and water-water-adjacent habitats. It resulted in the
formation of ornithocomplexes, similar to the communities of flood plains,
meadows, rivers and dumps. The population of water-water-adjacent cenoses is
then presented by nondivisible types. Their property is described on the
previous graph. Ruderal-town, settlement and dump type is divided into subtypes
of urban and rural building (8.1 and 8.2). The latter includes ornithocomplexes
of suburban gardens parks and public gardens. The common leader of these
subtypes is House and Tree Sparrow (Passer domesticus and P. montanus), while
the specific one is Rock Dove (Columba livia) and White Wagtail (Motacilla
alba). In towns the population density is somewhat higher than in the rural
buildings, parks and public gardens, but the background abundance is lower.
Ruderal communities form zonal-subzonal row, where the total bird abundance and
the background species richness constantly increased to the south. Therewith,
the population density here is considerable higher, than in all natural and
town, settlement and dump landscapes, while the number of the background
species is much lower. So it may be concluded that the main structural changes
in the bird population of the East-European plain on the level of type and
subtype are related to zonal heat supply, afforestation, degree of building and
ruderal capacity (based on the abundance of anthropogenic forages), swamping,
watering and differences in flowing capacity of reservoirs and rivers.
Similar trend in latitudinal changes from
tundra to steppe type is traced in the population of birds in the West Siberia.
But instead of one boreal type of the population this territory has two
independent types - the forest tundra and the forest type. Their intermediate
border coincides with that, intermediating the northern and middle taiga (Ravkin
et al., 1994). As in Europe, there is only weak threshold-overlapped connection
between the forest type and more northern type (in Europe - tundra and in
Siberia - the forest tundra). Maximum threshold-overlapped connection is
realized by the high bogy type of the community. Deviations from this trend in
the West-Siberian plain as well as in Europe are caused by the decreased
productivity of biocenoses in fields after the formation of monocultural
agrocenoses and by impoverished mineral supply in high bogs. In Siberia it
differs from the european trend by the higher mean abundance of birds in
Siberian marshes. Probably the european sample involves more bogs with
relatively low trophic capacity. The population of Siberian marshes enters the
meadow type of the communities and forms deviation from zonal row due to higher
productivity compared to the placor (zonal) vegetation formations. In Europe
the majority of meadows, even of the flood-plain meadows is ploughed up and has
poor population. The same is characteristic of the steppe type of the
population. The number of the relatively undeveloped steppes in Siberia, and
especially in Kazakhstan, is higher than in Europe. Thus, the average
population density of steppe type is higher and it doesn’t form, as in Europe,
deviations from the main trend.
In town, settlement, dump and ruderal
communities of the East-European plain zonal influence, though slightly limited
is also traced. This row is presented by only one type – ruderal-town,
settlement, dump. On the level of subtype it has obviously distinguished
population of dumps – tundra, northern taiga and southern taiga (more southern
dumps are not studied). The population density of dumps is higher than in the
remaining types of ornithocomplexes of unbuilt area (except for steppe
dubravas), Synanthropic types are leading here. In Siberia the influence of
zonality due to greater climate continentality is more pronounced. It resulted
in greater similarity of tundra settlement population with environs’
communities, and ornithocomplexes of the forest tundra villages form an
independent type, different from middle synanthropic type. The population of
studied taiga and the forest steppe dumps of West Siberia also forms separate
types with similar leading species inside, but in Europe it is added by House
Sparrow (Passer domesticus) and in Siberia forest tundra by White Wagtail
(Motacilla alba).
Not less pronounced is the zonality, traced
in water-water-adjacent communities in rivers and lakes (types 9-13). Regional
differences in types’ volume considerable associate with different
investigation intensity of their population. The data accumulated show that the
most frequently encountered leading species in Siberia are Ducks (Anatinae) and
in Europe Gulls (Larinae) and Stints (Charadrii).
Thus, spatial-typological structure of bird
population in the East-European and West-Siberian plains basically are similar
and determined by the same complement of environmental factors. But judging
from ornithocomplexes’ classification, the influence of ploughing, the
formation of monocultural agrocenoses, the formation age and the settlement
size in Europe is higher (Ravkin et al., 2001). In addition, the warming
Atlantic influence causes great, as compared with West Siberia, northwards
shift of the border between northern and middle supra-type groupings. In
Siberia hypoarctic species numerously penetrate the vast bogs and flood-plains
as well as light northern taiga forests, shifting the mentioned border far to
the south.
Even more pronounced and generalized
concepts on heterogeneity of ornithocomplexes are formed on the level of the
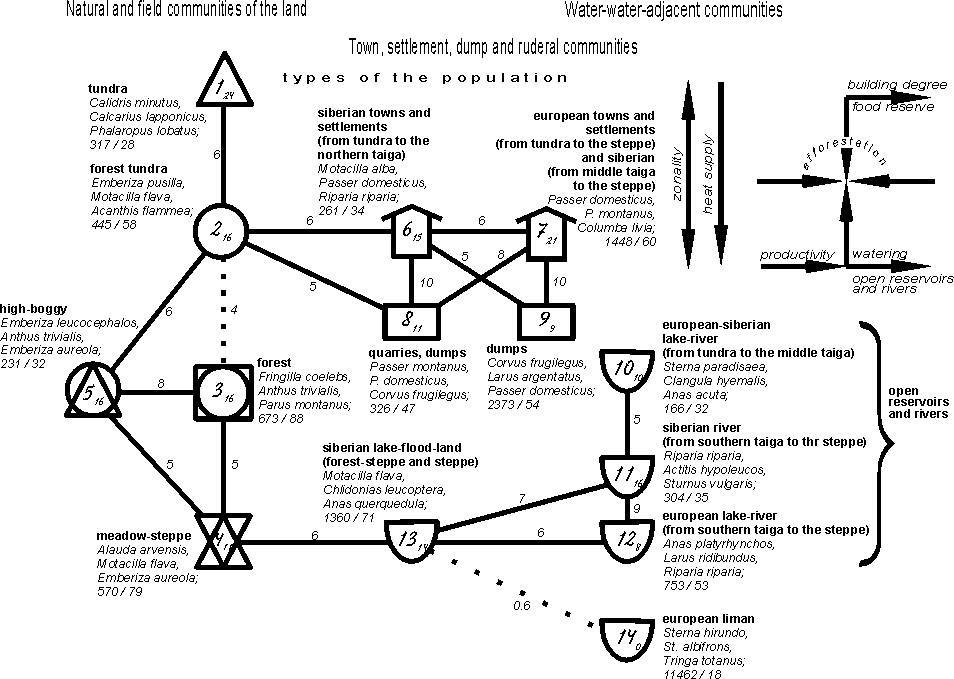
population type on the classification for both plains as a whole (fig. 3).
Central trend (types 1-4) shows zonal transition from the tundra type to the
forest-type, then through the high-boggy type of the population to the forest
and meadow-steppe type. In town, settlement, dump-ruderal part it was
traced the combined influence of
latitudinal and longitudinal change of the climate, where ornithocomplexes of
Siberian northern towns and villages considerable differ from the remaining
similar communities of Siberia and from all european communities (types 6 and
7). In ruderal part the population of quarries and dumps differ from that of
heaps (types 8 and 9). The population of northern-siberian town, settlement and
dump landscapes as well as of quarries and dumps is greatly associated with
ornithocomplexes of the forest-tundra type. Zonal differences are similarly
pronounced in the population of reservoirs and watersheds, but the provincial
differences are almost minimal. The population of water-water-adjacent habitats
is connected with that of the meadow by the communities of flood-lands and
grass-covered lakes.
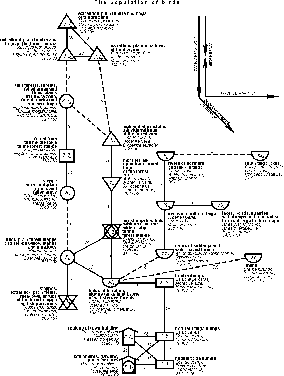
{kind=link}
Fig. 3. Spatial-typological
structure of bird communities of East-European and West-Siberian plains in the
first half of summer on the type level.
In contrast to structures, revealed for
each plain separately, the main zonal row of changes has only one shift as
high-bogy type, which as on the detailed graphs connects northern and middle
subtype groupings. The density and the background abundance of the population
in the main row constantly increases southwards up to the forest type,
decreasing in the meadow-steppe type. In the communities of high-bogy type the
sum abundance of birds is smaller than in the remaining types, except for the
population of northern lakes and rivers (from tundra to the middle taiga). The
number of the background species here is also lower, than in the majority of
the remaining communities. Only in ornithocomplexes of the tundra type
population and on limans the background species number is smaller, than in high
bogs, while in the same northern rivers and lakes their number is almost equal.
Differences in dominant bird species usually are traditional and show their
dependence on zonal phytocenoses' changes, properties of high bogs and town,
settlement and dump-rural landscapes. By and large this change coincides with
the previously described for separate structures. In the generalized classification
the dominant composition in water-water-adjacent communities changes the same
way as on the land, but somewhat differs from that in specific structures.
Gadwalls (Anatidae) and Arctic Tern (Sterna paradisaea) frequently dominate in
the north and Sand Martin (Riparia riparia), Common Sandpiper (Actitis
hypoleucos) and Starling (Sturnus vulgaris) – in the middle Siberia; Mallard
(Anas platyrhynchos), Black-heated Gull (Larus ridibundus) and Sand Martin
(Riparia riparia) – in Europe. Terns (Sterninae) and Redshank (Tringa totanus)
are leading in the Black sea limans.
By and large, the population trends are
determined by the heat supply (zonality), afforestation, productivity, food
reserve (especially in case of intensively urbanized areas and ruderality),
watering, flowing water and open reservoirs and rivers.
Conclusion
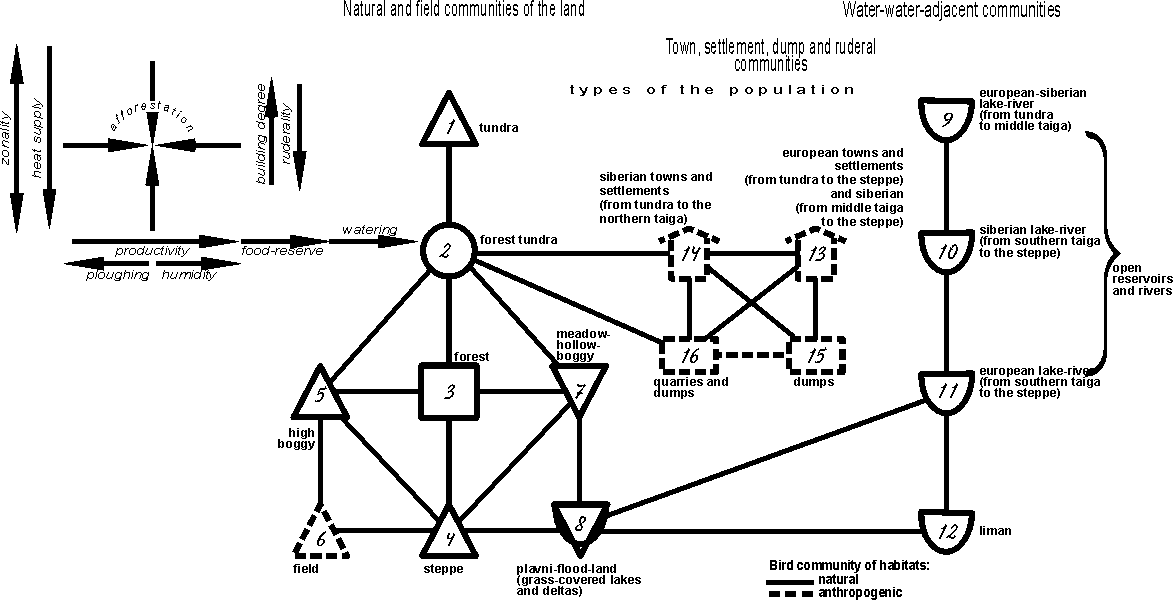
The comparison of the revealed structures
on each investigated plain and on the territory as a whole allowed to develop
the universal conceptual notion on the heterogeneity of the bird community
(fig. 4). The main zonal row of changes in natural and agricultural
communities is presented by the types of communities from tundra, via the
forest tundra and the forest, to the steppe type (1-4). It coincides with
trends of zonal changes in land vegetation. Shifts from this row are caused by
greatly different productivity of the communities, tending to natural decrease
on high bogs due to the impoverished mineral nutrition of phytocenoses and in
fields due to anthropogenic factors (types 5 and 6). The second row of shifts
is caused by natural increase of the productivity in meadow-marsh phytocenoses,
in plavni and flood-lands of grass-covered lakes and deltas (types 7 and 8).The
last type of the population causes changes in water-water-adjacent ornithocomplexes
(types 9-11), where zonality is equally pronounced, added by the provinciality
in the southern part, i.e. the lake-river type from tundra to the middle taiga
is unified, being divided southernmost into siberian and european variants. The
influence of zonality is also easily traced in town, settlement, dump and
ruderal communities. Ornithocomplexes of town, settlement and dump territories
considerable differ from the population of heaps on the one hand, and from the
quarries and dumps on the other, forming separate types (13 and 14; 15 and 16).
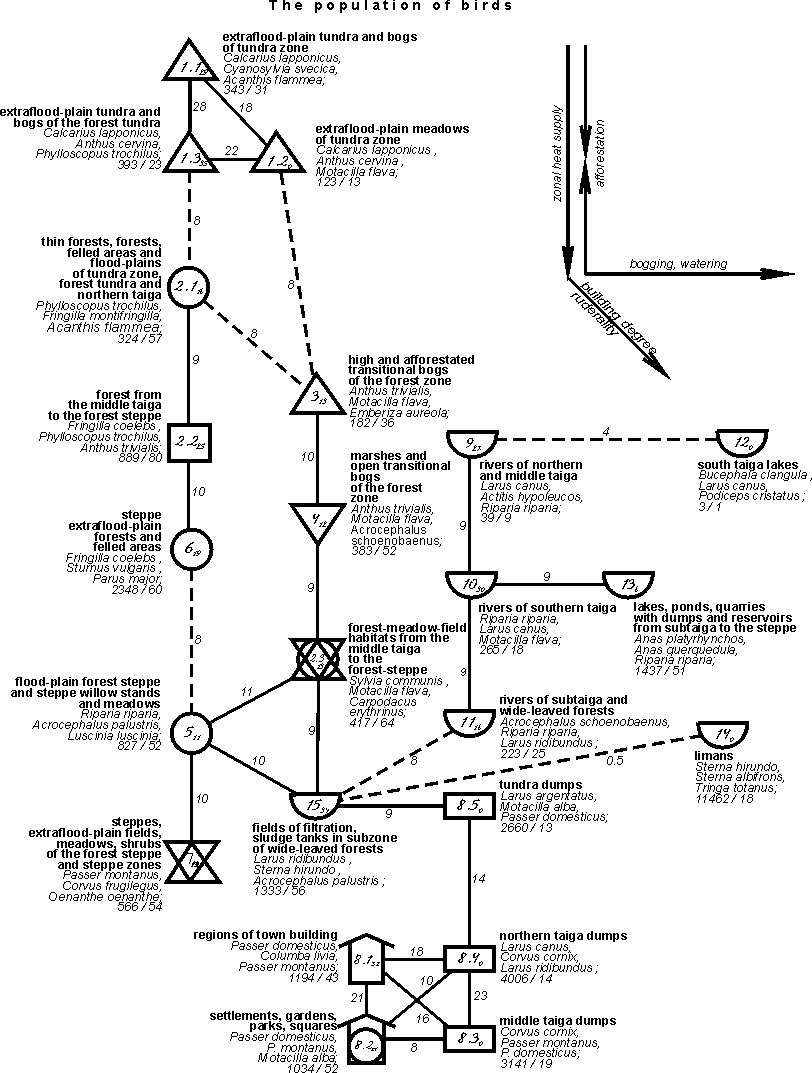
{kind=link}
Fig. 4. The final conceptual
population structure of birds of East-European and West-Siberian plains in the
first half of summer on the type level.
The informativity of structural notions,
given on the level of type for the East-European plain is 23% dispersion of
similarity coefficient and for the West Siberian plain and as a whole – by 38%.
Studies, basing this article were partially
financed by the Russian Foundation for Basic Research.
References
Ravkin E.S., Byshnev I.I., Kochanov S.K., et al.,
(2001) Spatial diversity on bird community of East-European and West-Siberian
plains. Advances and Problems of ornithology of Northern Eurasia at the end of
centuries, Kasan, "Magarif", pp. 212-236.
Ravkin E.S., Byshnev I.I., Kochanov S.K., et al.,
(2001) Spatial diversity on bird community of East-European and West-Siberian
plains. Advances and Problems of ornithology of Northern Eurasia at the end of
centuries, Kasan, "Magarif", pp. 212-236.