ПРОСТРАНСТВЕННО-ТИПОЛОГИЧЕСКАЯ
НЕОДНОРОДНОСТЬ НАСЕЛЕНИЯ ПТИЦ ВОСТОЧНО-ЕВРОПЕЙСКОЙ И
ЗАПАДНО-СИБИРСКОЙ РАВНИН
Е.С.Равкин*, Л.Г.Вартапетов, К.В.Торопов, Ю.С.Равкин, В.С.Жуков, И.В.Покровская, И.И.Бышнев, С.К.Кочанов
Центр эколого-географических разработок МосНПО “Радон”, Москва, 127562, Россия, ravkin_e@mtu-net.ru ; Лаборатория зоологического мониторинга ИСиЭЖ СО РАН, Новосибирск, 630091, Россия, zm@eco.nsc.ru ; Лаборатория биогеографии – ИГ РАН, Москва, 109117, Россия, pokrov@biogeo.msk.ru ; Березинский заповедник, Домжерицы, 211188, Беларусь; ИБ Коми НЦ УрО РАН, Сыктывкар, 167610, Россия.
* Адрес для переписки: Россия, 127562 Москва, Алтуфьевское шоссе 30-229.
Введение
Под пространственно-типологической структурой неоднородности населения птиц понимается общий характер территориальных различий орнитокомплексов, показанный на структурном графе, иллюстрирующем основные направления изменений сообществ птиц в зависимости от структурообразующих факторов среды.
Материалы
и методы
Для выявления пространственно-типологических структур населения птиц обеих равнин использованы материалы, накопленные в банке данных лаборатории зоологического мониторинга ИСиЭЖ СО РАН. Кроме сведений, собранных авторами, по Восточно-Европейской равнине использованы литературные источники (около 70) и данные вкладчиков банка, любезно предоставивших свои материалы для анализа. Все участники и районы работ перечислены в прежних публикациях [Равкин и др., 1994, 2000, 2001а]. Библиографическое описание использованных публикаций не приводится из-за ограниченности объёма статьи. В анализ включены результаты учетов птиц, проведенных в первой половине лета - с 16 мая по 15 июля 1880-1907, 1936-38, 1947-97 гг., хотя основная часть сведений собрана с конца 1970-х годов. Учёты проведены в основных по площади местообитаниях птиц всех природных зон Западно-Сибирской и Восточно-Европейской равнин (по физико-географическому районированию Атласа СССР [1986], т.е. исключая полупустыни и пустыни Прикаспийской низменности). Кроме того, в выборку включены материалы по населению птиц российской части Фенноскандии. Всего для расчетов использовано 2649 вариантов орнитокомплексов. Под вариантом населения птиц подразумевается повидовое обилие всех птиц, встреченных в данном местообитании за один сезон проведения учётов.
Повидовое обилие по вариантам населения усреднено в пределах выделов легенд карты растительности Европейской части СССР и Кавказа [1987] и Западной Сибири [1976] масштабов 1:2000000 и 1:1500000. В соответствии с этим, данные сгруппированы в 536 средних. Такое усреднение, до некоторой степени, нивелирует неравномерность в объемах собранного материала, а также годовые и индивидуальные отличия орнитокомплексов.
По рассчитанным так средним вычислены коэффициенты сходства Жаккара [Jaccard, 1902] для количественных признаков [Наумов, 1964]. Эти коэффициенты использованы для факторной классификации с поэтапной доразбивкой крупных классов, состоящих более, чем из пяти средних. Классификация проводилась с помощью одного из методов кластерного анализа [Трофимов, Равкин, 1980]. При этом для составления концептуальных классификационных схем использованы только интерпретируемые и понятные с предметной точки зрения разбиения. Пространственно-типологическая структура населения выявлялась методом корреляционных плеяд [Терентьев, 1959]. По матрице средних коэффициентов сходства орнитокомплексов, относящихся к таксонам классификации того или иного ранга, строился структурный граф. При этом, для построения структурного графа оставлялись связи выше порога, выбираемого так, чтобы граф наилучшим образом иллюстрировал выявленные тренды, и его можно было построить в двухмерном пространстве. Подробнее методологические и методические основы исследований опубликованы ранее [Равкин, Лукьянова, 1976; Равкин и др., 1994, 2001б]. Структурные представления о населении птиц Западной Сибири заимствованы из публикации Ю.С. Равкина с соавторами [1994].
Названия птиц даны по А.И. Иванову [1976].
Результаты
и обсуждение
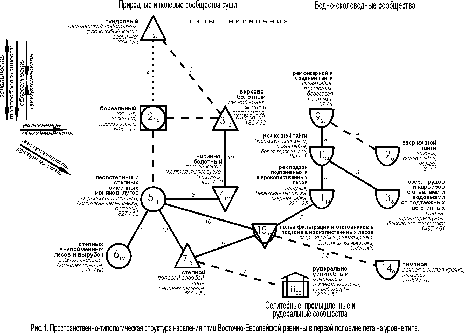
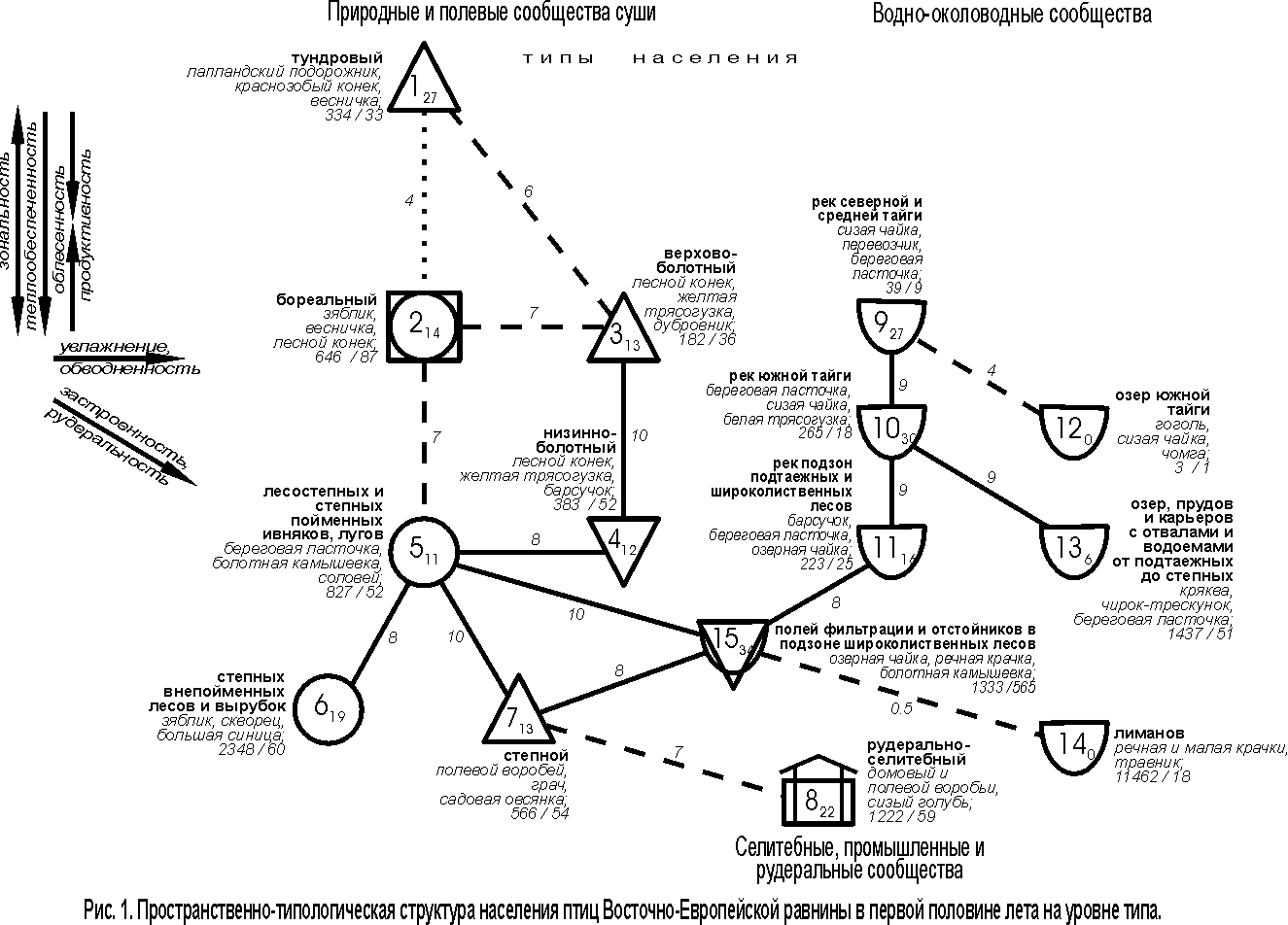
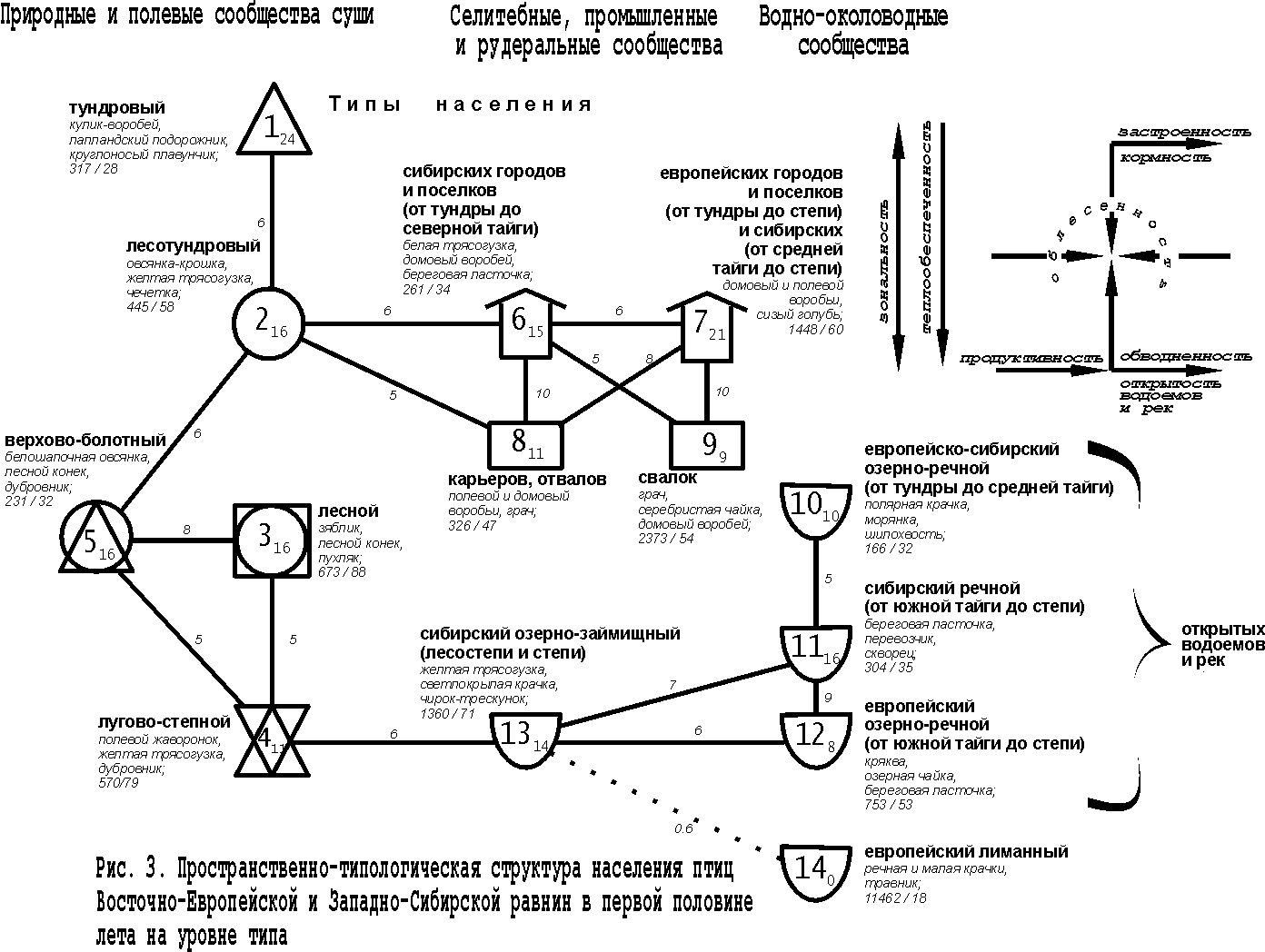
Пространственно-типологическая неоднородность животного населения может отражаться иерархической классификацией сообществ или структурным графом, т.е. классификацией со связями по значимому сходству между её таксонами того или иного уровня. Такая структура населения птиц Восточно-Европейской равнины на уровне типа сообществ отражает наличие трёх систем (зональных рядов). В первую из них входят природные и полевые сообщества, во вторую – селитебные, промышленные и рудеральные орнитокомплексы, в третью – водно-околоводные сообщества (рис. 1).
Рис.
1 – 4 построены в примерном обратном масштабе: чем выше межклассовое сходство
орнитокомплексов, тем ближе относительно друг друга расположены значки с номерами
типов или подтипов и индексами, обозначающими внутриклассовое сходство. Меж– и внутриклассовое сходство рассчитано как среднее от
коэффициентов сходства соответствующих вариантов населения. Нулевое сходство
соответствует единственному варианту в таксоне. Если связь невозможно отразить
в масштабе, она изображена пунктирной или штриховой линией и тоже имеет оценку
сходства. Квадратами обозначены классы с преобладанием
сообществ птиц лесов, кружками – тех местообитаний, где облесённые
участки чередуются с открытыми, а треугольниками – открытых местообитаний
(основанием вниз – с бедной продуктивностью, а вверх – с богатой), полуовалом –
водоёмов и рек, «домиком» – городов и посёлков, плоским прямоугольником –
свалок. Рядом со значком таксона приведено его название. На рис. 1 – 3
под названием таксона указаны первые по численности виды, плотность населения
птиц – особей/км2, и через косую черту – число
фоновых видов. Стрелками на всех рисунках показано увеличение проявления
основных структурообразующих факторов среды и определяемые ими тренды
населения.
{kind=link}
Первый ряд относится к первой из выделенных систем населения и представлен трендом по градиенту широтной теплообеспеченности (зональности) (типы 1, 2, 5, 6). По предметным соображениям он наиболее значим (первичен), хотя формально входящие в него типы лишь в одном случае связаны существенной связью (типы 5 и 6). Слабая запороговая связь прослеживается между бореальным и пойменным лесостепным типами (2 и 5), а тундровый и бореальный (1 и 2) имеют незначительное сходство и слабо связаны через население верховых болот. В реальном пространстве и в других системах населения чередование зон и соответствующих им типов сообществ сомнений не вызывает, поэтому предлагаемая трактовка предпочтительнее с концептуальной точки зрения.
Итак, тундровое населения птиц на Восточно-Европейской равнине сменяется бореальным, далее орнитокомплексами лесостепных и степных пойменных ивняков и лугов, затем степных внепойменных лесов и вырубок, а потом орнитокомплексами степного типа, положение которого двойственно. С одной стороны это, несомненно, зональный тип, с другой – за счёт обеднённого населения в полях, которое в него входит, он образует отклонение от основного ряда. За счёт этого тренд в нижней части графа раздваивается. В зональном ряду плотность населения максимальна в степных внепойменных лесах и на вырубках и убывает к северу, вплоть до тундрового типа. Число фоновых видов изменяется почти так же, как плотность, но наибольшие значения свойственны населению бореального типа. Это связано со значительной территорией, которую оно занимает.
Отклонение от основного широтного ряда составляет также население верхово-болотного и низинно-болотного типов. При заболачивании и большей или меньшей степени обеднения минерального питания фитоценозов их продуктивность уменьшается, что и определяет снижение плотности и фонового богатства населения. Смена в составе лидирующих видов при движении с севера на юг сводится к замене тундровых видов на лесные и опушечные или более южные, а в отклонениях и в селитебных, промышленных, рудеральных и водно-околоводных сообществах увеличивается доля болотных, луговых и водно-околоводных птиц, а также, по мере антропогенной трансформации исходных ландшафтов, синантропов.
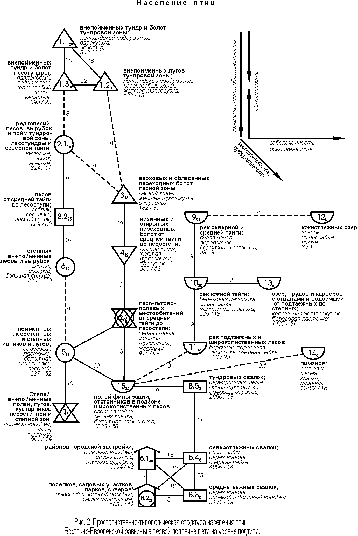
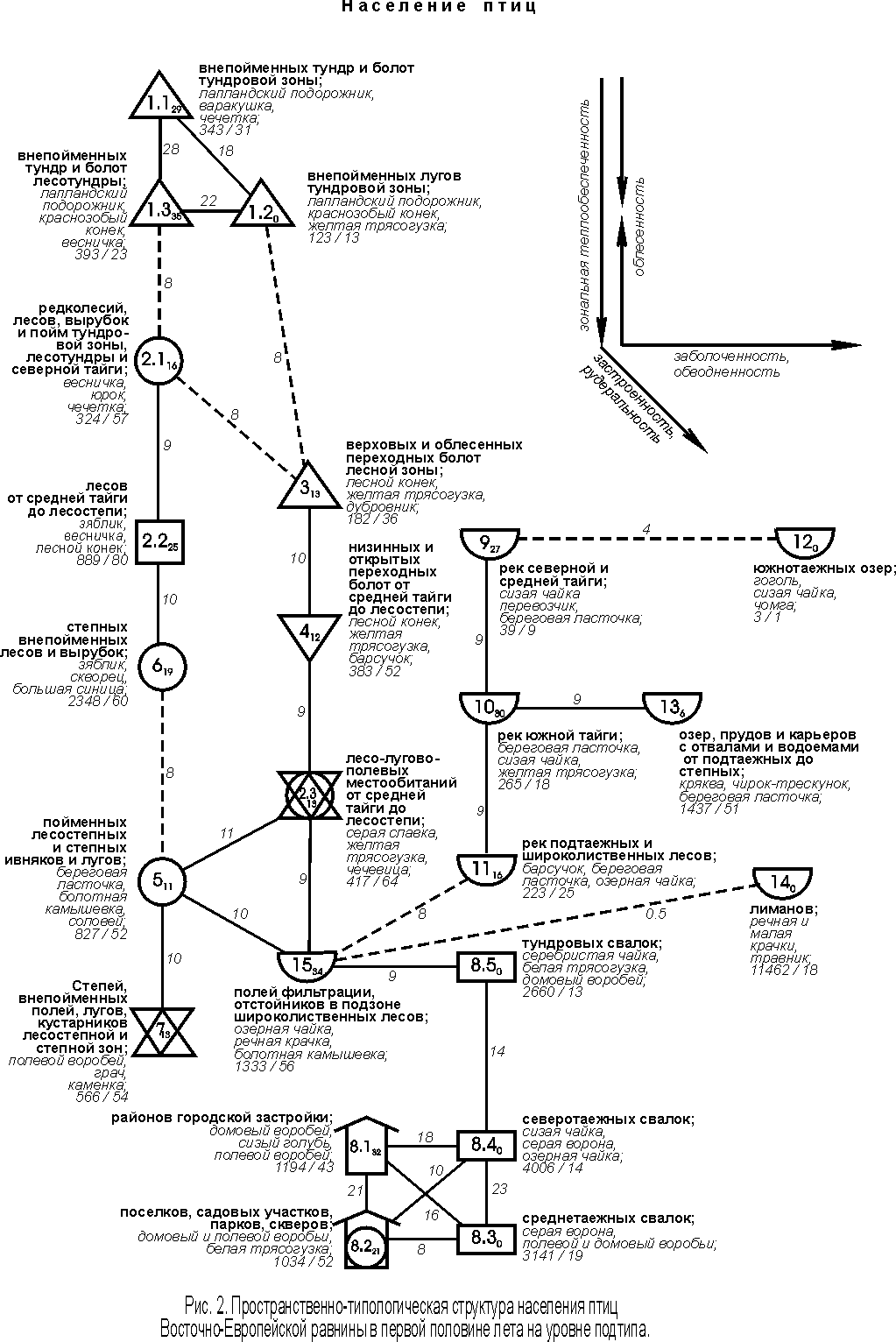
Структурный граф, построенный на уровне подтипа, в принципе подтверждает в подробностях то, что прослеживается на типовом уровне (рис. 2).
{kind=link}
Информативность подтиповых представлений почти вдвое выше (44% дисперсии коэффициентов сходства орнитокомплексов). Здесь основной зональный ряд изменений составляют орнитокомплексы внепойменных тундр и болот (сначала тундровой зоны, потом лесотундры). Далее следует население редколесий, лесов, вырубок и пойм тундровой зоны, лесотундры и северной тайги. Затем орнитокомплексы лесов (от средней тайги до лесостепи) сменяются населением степных внепойменных лесов и вырубок. За ними по сходству идут сообщества птиц пойменных лесостепных и степных ивняков в сочетании с лугами и, наконец, – внепойменных лугов, степей, полей и кустарников лесостепной и степной зон. При этом слабая связь прослеживается между населением лесотундровых болот и тундр (подтипы 1.1 и 1.3) и лесными орнитокомплексами (2.1, 2.2 и тип 6), а затем – между последними и сообществами ивняково-лугово-степного типов (5 и 7), которые не делятся на подтипы.
В зональном ряду подтипов населения суммарное обилие птиц почти неуклонно увеличивается к югу с 343 до 2348 особей/км2, а затем снижается в степном типе до 566 особей/км2. Исключение составляет плотность населения в редколесьях, лесах, вырубках и поймах тундровой зоны и лесотундры, которая несколько меньше ожидаемой. Обычно в равнинных северотаёжных лесах и горных редколесьях Сибири птиц несколько больше, чем в тундрах. Причина европейской специфики, видимо, во всё том же отепляющем влиянии Северо-Атлантического течения, которое в большей степени сказывается в приокеанических тундрах, чем в отдалённой от морского побережья лесотундре. Максимальным фоновым богатством отличается население лесных подтипов этого ряда (57 – 80 видов). Несколько меньше фоновых видов в самых южных открытых и закустаренных сообществах (52 – 54) и существенно меньше в орнитокомплексах тундрового типа (23 – 31). Состав лидеров в ряду сменяется от тундровых видов к лесотундровым и далее к лесным, пойменным и видам открытых лесостепных и степных местообитаний, что соответствует зональной смене растительности и определяется ею.
Параллельно описанному идёт также зональный ряд изменений в сообществах местообитаний, считающихся интразональными. В тундровой зоне и лесотундре это население внепойменных лугов (подтип 1.2), в лесной зоне и лесостепи – болот (типы 3 и 4) и лесо-лугово-полевых местообитаний (подтип 2.3). В этих типах и подтипах населения суммарное обилие птиц всегда ниже, чем в местообитаниях основного зонального ряда, что, возможно, связано с уменьшением продуктивности фитоценозов. В лесной зоне это верховые, переходные и близкие к ним низинные болота, трофность которых ниже, чем в лесах и богатых по трофности низинных болотах и лугах, особенно пойменных. В лесо-лугово-полевых местообитаниях кормность ниже из-за создания монокультурных агроценозов и, возможно, из-за интенсивного сенокошения и перевыпаса скота в лугах. В Сибири сохранность лугов из-за менее интенсивного ведения сельского хозяйства выше, а обследованные низинные болота, особенно пойменные, богаче по трофности и больше привлекают птиц. Поэтому на Западно-Сибирской равнине лугово-болотные варианты населения имеют высокую плотность и отклоняются от основного ряда по причине большей, а не меньшей (как в Европе) продуктивности. Состав лидеров в ряду отклонений также совпадает с зональной сменой растительности. Лугово-тундровые птицы сменяются болотно-луговыми и видами мозаичных насаждений и кустарников. Плотность населения и богатство фоновых видов в этом ряду неуклонно увеличивается с севера на юг.
Через население птиц полей фильтрации и отстойников этот ряд связан с селитебно-рудеральными и водно-околоводными сообществами. Отстойники и поля фильтрации из-за высокой трофности привлекают много птиц, как лугово-болотных, так и водно-околоводных. За счёт этого формирующиеся здесь орнитокомплексы близки к сообществам пойм, лугов, рек и свалок. Население водно-околоводных ценозов представлено далее не делящимися типами. Их специфика описана по предыдущему графу.
Рудерально-селитебный тип делится на подтипы городской и сельской застройки (8.1 и 8.2). Последний из них включает ещё и орнитокомплексы пригородных садово-огородных участков, парков и скверов. Общими лидерами для этих подтипов являются домовый и полевой воробьи, а специфичными – сизый голубь и белая трясогузка. В городах плотность населения несколько выше, чем в участках сельской застройки, а также в парках и скверах; фоновое богатство, напротив, меньше. Рудеральные сообщества образуют зонально-подзональный ряд, в котором общее обилие птиц и богатство фоновых видов неуклонно увеличиваются к югу. При этом плотность населения здесь значительно выше, чем во всех естественных и селитебных ландшафтах, а число фоновых видов существенно ниже.
В ряду зонально-подзональной смены орнитокомплексов суши, начиная от средней тайги, к югу отмечен устойчивый инверсионный характер изменения плотности и фонового богатства орнитокомплексов (показатели уменьшаются при движении с севера на юг). Это определяется увеличением роли антропогенных изменений среды обитания птиц, в основном влияния распашки, достигающей максимума в степной зоне (Равкин и др, 2001а).
В итоге можно констатировать, что основные структурные изменения в населении птиц Восточно-Европейской равнины на уровне типа и подтипа связаны с зональной теплообеспеченностью, облесённостью, застроенностью и рудеральностью (через обилие антропогенных кормов), распашкой, а также с заболоченностью, обводнённостью и различием в проточности водоёмов и рек.
Сходный тренд широтных изменений от тундрового до степного типа прослеживается и в населении птиц Западной Сибири. Только на этой территории вместо единого бореального типа населения имеется два самостоятельных – лесотундровый и лесной. Граница между ними совпадает с границей между северной и средней тайгой [Равкин и др., 1994]. Так же, как в Европе имеется лишь слабая запороговая связь между лесным типом и лежащим севернее (в Европе тундровым, в Сибири лесотундровым). Максимальная запороговая связь идёт через верхово-болотный тип населения. Отклонения от тренда на Западно-Сибирской равнине, так же, как в Европе, связаны с уменьшением продуктивности биоценозов в полях после создания монокультурных агроценозов и в связи с обеднением минерального питания на верховых болотах. Отличия от европейского тренда заключаются в том, что население сибирских низинных болот в среднем богаче, чем в Европе. Видимо в выборке по Европе больше болот с относительно низкой трофностью. Население сибирских низинных болот входит в луговой тип сообществ и образует отклонение от зонального ряда в связи с большей продуктивностью по сравнению с плакорными (зональными) растительными формациями. В Европе подавляющая часть лугов, даже пойменных, распахана и имеет бедное население. То же можно сказать и о степном типе населения. Относительно нетронутых степей в Сибири, и особенно в Казахстане, больше, чем в Европе. Соответственно плотность населения в среднем по степному типу выше и он не образует, как в Европе, отклонения от основного тренда.
В селитебных, промышленных и рудеральных сообществах Восточно-Европейской равнины также прослеживается влияние зональности, хотя и в несколько усечённом виде. Этот ряд представлен всего одним типом – рудерально-селитебным. На уровне подтипа в нём чётко выделяется население свалок – тундровых, северотаёжных и южнотаёжных (более южные свалки не обследованы). Плотность населения на свалках выше, чем в остальных типах орнитокомплексов незастроенной суши (за исключением степных дубрав). Лидируют здесь синантропы. В Сибири влияние зональности из-за большей континентальности климата проявляется чётче. В результате население тундровых посёлков более сходно с окрестными сообществами, а орнитокомплексы лесотундровых сел образуют самостоятельный тип, отличный от срединного синантропного. Население обследованных в Западной Сибири таёжных и лесостепных свалок тоже образует отдельные типы. Лидирующие виды в них сходны, но в Европе в их число входит домовый воробей, а в сибирской лесотундре – белая трясогузка.
Не менее чётко зональность прослеживается и в водно-околоводных сообществах как на реках, так и на озёрах (типы 9 – 13). Региональные отличия в объёме типов в значительной степени связаны с разной обследованностью их населения. По собранным данным прослеживается, что в качестве лидеров в Сибири, особенно в северной её части, чаще встречаются утки, а в Европе – чайки и кулики.
Итак, пространственно-типологические структуры населения птиц Восточно-Европейской и Западно-Сибирской равнин принципиально одинаковы и определяются одним и тем же набором факторов среды. Однако, судя по классификации орнитокомплексов, влияние распашки и создания монокультурных агроценозов, возраста формирования и размера населённых пунктов в Европе выше [Равкин и др. 2000, 2001б]. Кроме того, отепляющее влияние Атлантики на этой территории приводит к значительному, по сравнению с Западной Сибирью, смещению к северу границы между северной и срединной надтиповыми группировками. В Сибири гипоарктические виды в значительном количестве проникают по обширным болотам и поймам, а также по разреженным северотаёжным лесам, сдвигая указанную выше границу далеко на юг.
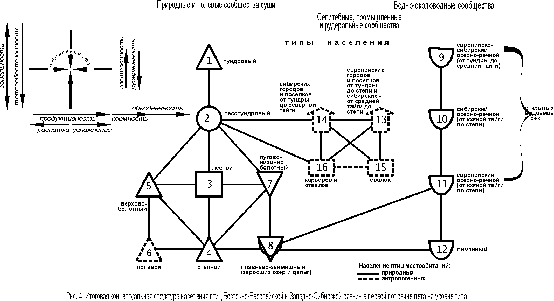
Ещё более чёткие и обобщённые представления о неоднородности орнитокомплексов формируются на уровне типа населения по классификации, составленной по обеим равнинам в целом (рис. 3).
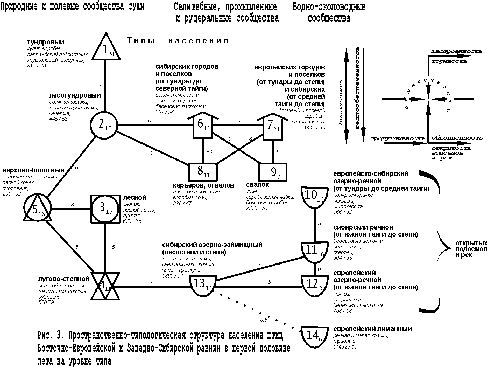
{kind=link}
Центральный тренд (типы 1 – 4) иллюстрирует зональный переход от тундрового типа к лесотундровому, далее через верхово-болотный тип населения к лесному и лугово-степному. В селитебно-рудеральной части прослеживается объединённое воздействие широтного и долготного изменения климата – орнитокомплексы сибирских северных городов и посёлков существенно отличаются от остальных аналогичных сообществ Сибири и всех европейских (типы 6 и 7). В рудеральной части население карьеров и отвалов отличается от сообществ свалок (типы 8 и 9). Население северо-сибирских селитебных ландшафтов, а также карьеров и отвалов значимо связано с орнитокомплексами лесотундрового типа. В населении водоёмов и водотоков не менее определённо прослеживаются зональные отличия и почти не сказываются провинциальные. Через сообщества займищ и зарастающих озёр население водно-околоводных местообитаний связано с луговым.
В отличие от структур, выявленных для каждой из этих равнин в отдельности, основной зональный ряд изменений имеет лишь одно отклонение в виде верхово-болотного типа, который, так же, как на частных графах, связывает северную и срединную надтиповые группировки. Плотность и фоновое богатство населения в основном ряду неуклонно возрастают к югу вплоть до лесного типа, а затем уменьшаются в лугово-степном. В сообществах верхово-болотного типа суммарное обилие птиц меньше, чем в остальных типах, кроме населения северных озёр и рек (от тундры до средней тайги). Число фоновых видов здесь тоже ниже, чем в большинстве остальных сообществ. Только в орнитокомплексах тундрового типа населения и на лиманах фоновых видов меньше, чем на верховых болотах, а на тех же северных реках и озёрах их примерно одинаковое количество. Отличия в лидирующих видах птиц в целом традиционны и иллюстрируют зависимость их от смены зональных фитоценозов, специфики верховых болот и селитебно-рудеральных ландшафтов. Эта смена в целом совпадает с ранее описанной по отдельным структурам. В единой классификации в водно-околоводных сообществах состав лидеров изменяется так же, как на суше, но несколько отличается от таковых на частных структурах. Так на севере нередко лидируют утки и полярная крачка, в срединной части Сибири – береговая ласточка, перевозчик и скворец, а в Европе – кряква, озёрная чайка и береговая ласточка. На черноморских лиманах это крачки и травник.
В целом, тренды в населении определяются теплообеспеченностью (зональностью), облесённостью, продуктивностью, кормностью (особенно при высокой застроенности и рудеральности), а также обводнённостью, проточностью и открытостью водоёмов и рек.
Заключение
Сопоставляя все выявленные структуры отдельно по исследуемым равнинам и по их территории в целом, можно сформировать единые концептуальные представления о неоднородности населения птиц (рис. 4).
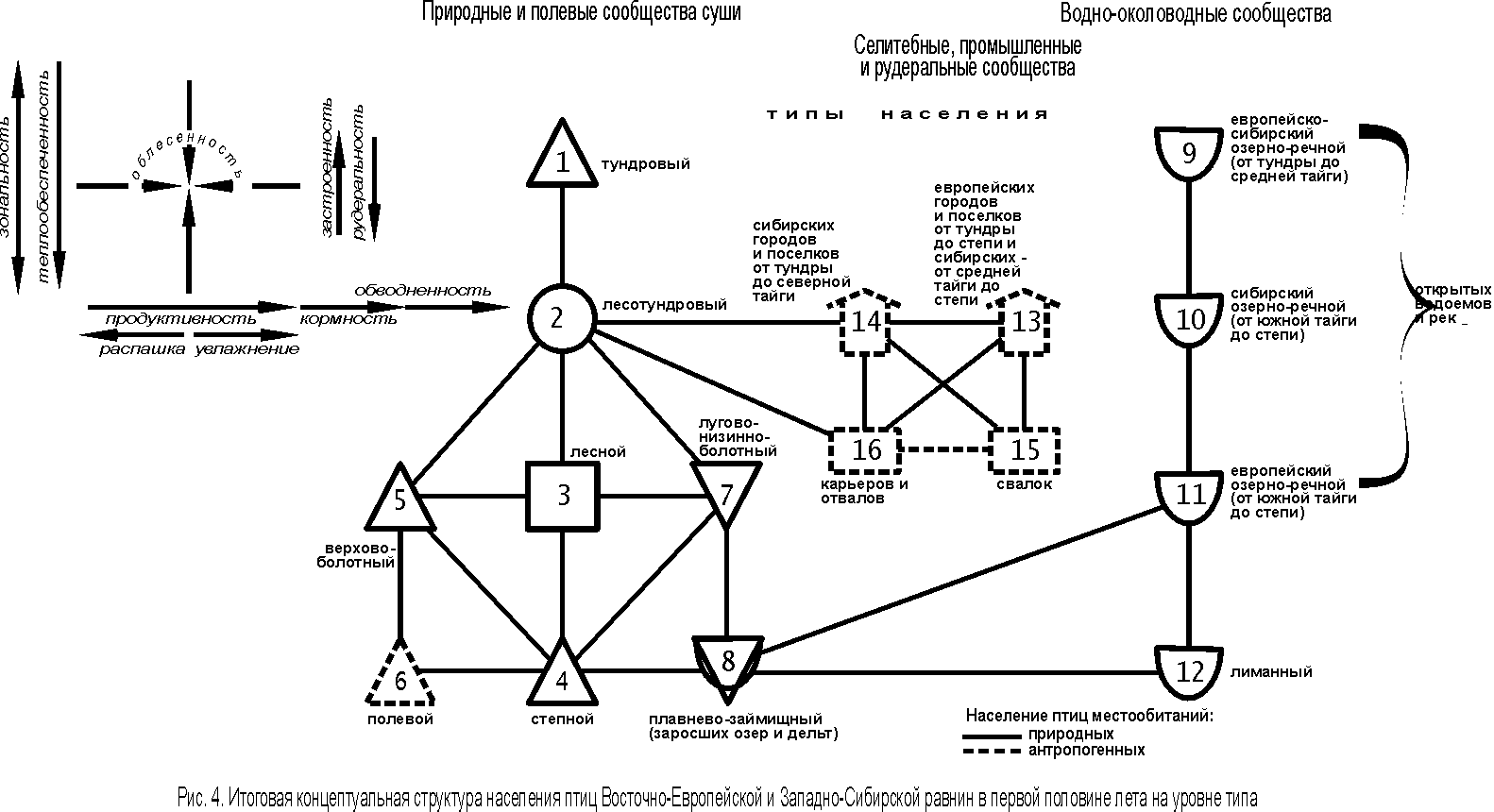{kind=link}
Основной зональный ряд изменений в природных и полевых сообществах представлен типами населения от тундрового, через лесотундровый и лесной, к степному (1 – 4). Он совпадает с тенденцией зональных изменений в растительности суши. Отклонения от этого ряда определяются существенно отличающейся продуктивностью сообществ, в сторону уменьшения по естественным причинам на верховых болотах, в связи с обеднением минерального питания фитоценозов, и антропогенным - в полях (типы 5 и 6). Второй ряд отклонений связан с естественным увеличением продуктивности в лугово-низинно-болотных фитоценозах, а также в плавнях и займищах заросших озёр и дельт (типы 7 и 8). Через последний тип населения изменения идут к водно-околоводным орнитокомплексам (типы 9 – 12), где столь же чётко прослеживается зональность и, кроме того, в южной части ещё и провинциальность (региональность), т.е. озёрно-речной тип от тундры до средней тайги един, а южнее делится на сибирский и европейский варианты. Влияние региональности легко проследить и в селитебных, промышленных и рудеральных сообществах. Орнитокомплексы селитебных территорий существенно отличаются от населения свалок с одной стороны и карьеров, отвалов с другой, и образуют отдельные типы (13 и 14; 15 и 16).
Информативность структурных представлений на уровне типа по Восточно-Европейской равнине составляет 23% дисперсии матрицы коэффициентов сходства орнитокомплексов, по Западно-Сибирской равнине и в целом – по 38%.
Благодарности
Исследования, послужившие основой для настоящей статьи, частично финансировались РФФИ.
Литература
Атлас
СССР. ГУГиК при СМ СССР, 1986. С. 120.
Иванов
А.И. Каталог птиц Советского Союза. – Л.: Наука, 1976, -276 с.
Наумов
Р.Л. Птицы в очагах клещевого энцефалита Красноярского края // Автореф. дис. ... канд. биол.
наук. – М., 1964, - 19 с.
Равкин Е.С., Бышнев И.И., Кочанов С.К. и др. Пространственная неоднородность летнего населения птиц Восточно-Европейской и Западно-Сибирской равнин // Экология и рациональное природопользование на рубеже веков. Итоги и перспективы. Мат-лы международной конф. Т. 1. – Томск, 2000. С. 163-164.
Равкин Е.С., Бышнев И.И., Кочанов С.К. Пространственное разнообразие летнего населения птиц Восточно-Европейской и Западно-Сибирской равнин // Достижения и проблемы орнитологии Северной Евразии на рубеже веков (Труды международной конференции «Актуальные проблемы изучения и охраны птиц Восточной Европы и Северной Азии). Казань, 2001а. С.212–236.
Равкин Е.С., Равкин Ю.С., Вартапетов Л.Г. и др. Классификация летнего населения птиц равнин Северной Евразии // Сибирский экологический журн., 2001б, - с. 741-766.
Равкин Ю.С., Лукьянова И.В. География позвоночных южной тайги Западной Сибири. – Новосибирск: Наука, 1976, - 360 с.
Равкин Ю.С., Вартапетов Л.Г., Юдкин В.А. и др. Пространственно-типологическая структура и организация летнего населения птиц Западно-Сибирской равнины // Сибирский экологический журн., 1994, 4, - с. 303-320.
Терентьев П.В. Метод корреляционных плеяд // Вестник Ленинградского университета, 1959, № 9, - с. 137-141.
Трофимов
В.А., Равкин Ю.С. Экспресс-метод оценки
связи пространственной неоднородности животного населения и факторов
среды // Количественные методы в экологии животных. - Л, 1980, - с. 113-115.
Jaccard P. Lois de distribution
florale dans la zone
alpine. –Bull. Soc. Vaund. Sci. Nat. 1902, vol. 38:
- p. 69-130.
SUMMARY
SPATIAL-TYPOLOGICAL
HETEROGENEITY OF BIRD COMMUNITY OF EAST-EUROPEAN AND WEST-SIBERIAN PLAINS
E.S. Ravkin*1, L.G. Vartapetov2,
K.V. Toropov2, Yu.S. Ravkin2,
S.M. Tsybulin2, V.S. Zhukov2,
I.V. Pokrovskaja3, I.I. Byshnev4.
S.K. Kochanov5
1. Center of ecologic-geographic developments SIA “Radon”, Moscow, 127562, Russia, ravkin_e@mtu-net.ru
;3. Laboratory
of biogeography – IG RAS, Moscow, 109117, Russia, pokrov@biogeo.msk.ru;
4. Berezinsky reserve, Domzheritzy,
211188, Byelorussia;
5. NB Komi NC UrORAS, Syktyvkar, 167610, Russia
*To whom
correspondence should be addressed. Russia, 127562
Moscow, Altufievskoye Road 30-229