ПРОСТРАНСТВЕННО-ТИПОЛОГИЧЕСКАЯ СТРУКТУРА И ОРГАНИЗАЦИЯ НАСЕЛЕНИЯ НАЗЕМНЫХ ПОЗВОНОЧНЫХ ЗАПАДНОЙ СИБИРИ (земноводные, птицы и мелкие млекопитающие)
Ю.С.Равкин, Л.Г.Вартапетов, В.А.Юдкин, И.В.Покровская, К.В.Торопов,
В.С.Жуков, С.М.Цыбулин, А.К.Добротворский, Б.Н.Фомин,
В.В.Панов, В.Н.Блинов, Т.К.Блинова, А.М.Адам,
И.Н.Богомолова, Е.Л.Шор, В.П.Стариков, С.А.Соловьёв, В.М.Ануфриев, А.А.Ананин, Г.М.Тертицкий
Институт систематики и экологии животных СО РАН,
Россия, 630091, Новосибирск,
ул. Фрунзе 11
АННОТАЦИЯ
Описаны пространственные изменения в населении наземных позвоночных Западной Сибири на примере земноводных, птиц и мелких млекопитающих в целом, рассмотренных по их численности и количеству трансформируемой энергии. Оценена сила и общность связи неоднородности сообществ и среды. Показано несовпадение геоботанических и ландшафтных границ и территориальной неоднородности различных таксоценов животных.
ВВЕДЕНИЕ
Пространственно-типологическая структура и организация
населения земноводных, птиц и мелких млекопитающих Западно-Сибирской равнины
описаны сравнительно полно по каждой группе этих животных отдельно и, кроме
того, проведены соответствующие сопоставления [1–8]. В
настоящем сообщении даётся общий анализ неоднородности сообществ
всех трёх групп животных вместе. На их долю приходится большая часть
населения наземных позвоночных, поэтому есть основание предполагать, что общий
характер изменений основных показателей этих таксоценов
и связи неоднородности их сообществ с факторами среды будут аналогичными с таковыми
по всем наземным позвоночным исследуемой территории. Районы работ и годы
проведения учётов описаны в упомянутых публикациях, так же как и полный список
лиц, собиравших материал, который послужил основой для данного сообщения. В нём
приводится анализ результатов учётов, проведённых только во второй половине
лета (с 16.07 по 31.08).
Для сравнимости результаты относительных учётов земноводных и мелких млекопитающих приведены к абсолютным значениям ранее описанным способом [9]. Приблизительность подобного приведения оговорена там же. После этого все материалы раздельно по группам животных усреднены по выделам карты «Растительность Западно-Сибирской равнины» [10] с учётом подзональной принадлежности и дифференцированно по трём примерно равным долготным полосам. Последнее сделано для того, чтобы при анализе не потерять информацию о провинциальных отличиях населения, если таковые окажутся значимыми. После этого, опять же отдельно по долготным полосам и выделам указанной карты, объединены варианты населения всех трёх групп животных. Кроме этих вариантов так же усреднены и объединены материалы по населению посёлков и водно-околоводных сообществ. В последних условно принято, что земноводные, закончившие метаморфоз, и мелкие млекопитающие не встречаются на акваториях и попадают в пространство, занимаемое этими комплексами лишь по сплавинам и в узкой прибрежной полосе, временно освобождающейся от воды при обмелении и отступлении приливно-нагонных вод. Остальные допущения и методы исследования оговорены в указанных ранее публикациях [1-9].
В
результате описанного усреднения и объединения исходных данных получено 363 средних
варианта населения. По этим средним рассчитаны коэффициенты сходства Жаккара [11] для количественных признаков [12]. По ним
проведена классификация населения с помощью одного из методов кластерного
анализа [13]. Анализ выделившихся при этом классов, доразделённых
на подклассы и более мелкие подгруппы, показал, что влияние провинциальности не
значимо по сравнению с другими более сильными факторами среды. После этого
данные вновь усреднены по выделам той же карты уже независимо от принадлежности
к той или иной долготной полосе. В результате этого усреднения получен 271
вариант населения. На этих средних вновь выполнена классификация сообществ. Для
типов и подтипов населения рассчитаны внутри- и межгрупповые связи и на их
основе выявлены основные изменения населения (тренды) и построены структурные
графы.
С
помощью линейной качественной аппроксимации (подробнее
смотри упомянутые публикации и ссылки в них) определена сила и общность связи
неоднородности населения с факторами среды и рассчитана информативность
структурных и классификационных представлений. Эти расчёты выполнены на матрице
коэффициентов сходства 363 усреднённых вариантов населения, что позволило
оценить силу и общность связи и с провинциальными отличиями. Все оценки связи
выполнены на 100 вариантах, отобранных из 363 с помощью генератора случайных
чисел с учётом соотношения площадей, занимаемых различными местообитаниями.
Такие расчёты проведены 8 раз, что позволило оценить ошибку выборочности и
доверительные интервалы.
Весь
комплекс обработки выполнен по показателям обилия животных и по количеству
трансформируемой ими энергии [9]. Последнее связано с тем, что учёты земноводных
проводились вскоре или во время выхода молодых из водоёмов выплода. Доля
сеголеток в населении в это время очень велика, а распределение в значительной
мере зависит не от общих ландшафтных условий местности, различиями в которых мы
и объясняем неоднородность населения, а определяется близостью ловчих канавок к
мелким водоёмам с успешным выплодом.
Информацию о них нельзя считать с природно-географических карт и
аэрокосмических снимков. Поэтому полнота объяснения причин
неоднородности населения сравнительно невелика. Роль сеголеток в
биоценозе значительно меньше, чем молодых и взрослых особей земноводных, не
говоря уже о более крупных животных – млекопитающих и птицах, которые выполняют
больший объём биоценотической работы ещё и за счёт более интенсивного обмена.
Поэтому количество энергии, трансформируемой животными, ближе к
биоценотическому значению и отражает территориальные отличия населения в более общем виде, чем по обилию [14].
КЛАССИФИКАЦИЯ
ЖИВОТНОГО НАСЕЛЕНИЯ ПО
ПОКАЗАТЕЛЯМ
ЧИСЛЕННОСТИ
А. Северная
надтиповая группировка
1. Арктический тундровый тип
населения (подзоны арктических тундр; лидируют сибирский лемминг,
кулик-воробей, чернозобик, круглоносый плавунчик, турухтан; плотность населения
906 особей/км2; общее число встреченных
видов 40 / в том числе фоновых – 29; доля представителей арктического типа
фауны 98%)[1].
Подтипы населения:
1.1 – тундр, пойменных ивняков,
лугов (сибирский лемминг, кулик-воробей, рогатый жаворонок, лапландский
подорожник, турухтан, 850; 29/19; арктического типа 99%),
1.2 – болот (кулик-воробей,
чернозобик, круглоносый плавунчик, белохвостый песочник, сибирский лемминг;
1439; 29/25; арктического типа 98%),
1.3 – тампов
(сибирский лемминг, турухтан, белохвостый песочник, кулик-воробей, круглоносый
плавунчик; 538; 25/21; арктического типа 92%).
2. Субарктический тундровый тип
населения (подзоны субарктических тундр, тундр и бугристых болот в
пределах лесотундры и северной тайги;сибирский
лемминг, тундряная бурозубка, полёвка-экономка, средняя бурозубка, остромордая
лягушка; 3275; 119/44; арктического типа 42%, тундро-лесостепных реликтов 32%,
транспалеарктов 14%).
Подтипы
населения:
2.1 – северных субарктических
тундр, болот и лугов (сибирский лемминг, тундряная бурозубка, краснозобый
конёк, лапландский подорожник, копытный лемминг; 6485; 64/33; арктического типа
76%, тундро-лесостепных реликтов 21%);
2.2 – южных субарктических низкокустарниковых тундр (сибирский лемминг, тундряная
бурозубка, полёвка-экономка, лапландский подорожник, копытный лемминг; 887;
63/24; арктического типа 45%, тундро-лесостепных реликтов 25%, транспалеарктов
18%);
2.3 – южных субарктических
кустарниковых тундр (полёвка-экономка, тундряная бурозубка,
чечётка, сибирский лемминг, средняя бурозубка; 2317; 62/34; транспалеарктов
45%, тундро-лесостепных реликтов 26%, арктического типа 15%);
2.4 – южных субарктических тундровых бугристых и полигональных болот (тундряная и
средняя бурозубки, сибирский лемминг, краснозобый конёк, лапландский
подорожник; 1248; 55/30; тундро-лесостепных реликтов 58%, транспалеарктов 18%,
арктического типа 16%);
2.5 – лесотундровых и северотаёжных тундр и бугристых болот (остромордая лягушка,
тундряная бурозубка, средняя бурозубка, полёвки – красная и экономка; 1181;
92/35; европейского типа 42%, тундро-лесостепных реликтов 20%, транспалеарктов
17%, сибирского типа 11%);
2.6 – субарктических пойм
крупных рек (тундряная и средняя бурозубки, полёвка-экономка, краснозобый конёк, сибирский лемминг; 8159; 62/33;
тундро-лесостепных реликтов 59%, транспалеарктов 26%).
Б.
Срединная надтиповая группировка.[2]
3. Лесотундровый тип населения
(редколесий, редкостойных и сосновых лесов и верховых
болот в пределах лесотундры и северной тайги; остромордая лягушка, красная
полёвка, средняя и тундряная бурозубки, тёмная полевка; 4507; 167/58;
сибирского типа 34%, транспалеарктов 28%, европейского и арктического типов по
14%).
3.1. Подтип населения редколесий
и редкостойных лесов (остромордая лягушка, красная полёвка, средняя и тундряная
бурозубки, полёвка-экономка; 4499; 126/46; европейского типа 32%, сибирского
30%, транспалеарктов 24%; тундро-лесостепных реликтов 13%).
Классы
населения:
3.1.1 – лесотундры (тундряная бурозубка, красная полёвка, средняя бурозубка,
овсянка-крошка, чечётка; 2550; 95/37; сибирского типа 36%, тундро-лесостепных
реликтов 28%, транспалеарктов 26%);
3.1.2 – северной тайги
(остромордая лягушка, красная полёвка, средняя и тундряная бурозубки,
полёвка-экономка; 6015; 106/45; европейского типа 40%, сибирского 27%,
транспалеарктов 24%).
3.2. Подтип населения северотаёжных сосновых лесов и верховых болот (остромордая
лягушка, средняя бурозубка, красная полёвка, тундряная
бурозубка, тёмная полёвка; 4522; 144/62; европейского типа 41%, сибирского и
транспалеарктов по 25%).
4. Лесной тип
населения (лесов, кроме редкостойных и
сосновых в пределах лесотундры и северной тайги; пойм крупных рек от лесотундры
до лесостепи; внепойменных низинных и верховых болот,
полей, перелесков, лугов и степей от средней тайги до лесостепи; займищ;
лесостепных свалок; остромордая лягушка, серая жаба, обыкновенная бурозубка,
сибирский углозуб, средняя бурозубка; 50392; 334/128;
европейского типа 88%).
Подтипы
населения:
4.1 – темнохвойных, смешанных и
мелколиственных нередкостойных лесов лесотундры и
северной тайги (остромордая лягушка, красная полёвка, средняя, тундряная и обыкновенная
бурозубки; 13632; 141/66; европейского типа 56%, сибирского 19%, транспалеарктов
16%);
4.2 – пойм крупных рек в
пределах лесотундры и северной тайги (остромордая лягушка, обыкновенная
бурозубка, полёвка-экономка, сибирский углозуб,
тундряная бурозубка; 22165; 139/70; европейского типа 86%);
4.3 – темнохвойных,
темнохвойно-мелколиственных и мелколиственных лесов, гарей, вырубок,
полей-перелесков средней и южной тайги (остромордая лягушка, серая жаба,
обыкновенная и средняя бурозубки, красная полёвка; 32145; 230/109; европейского
типа 70%, сибирского 15%, транспалеарктов 15%).
Классы
населения:
4.3.1 – средней тайги (серая жаба,
остромордая лягушка, средняя и обыкновенная бурозубки, красная полёвка; 29763;
155/73; европейского типа 67%, сибирского 16%, транспалеарктов 15%),
4.3.2 – южной тайги (остромордая
лягушка, серая жаба, обыкновенная и средняя бурозубки, красная полёвка; 33244; 219/115; европейского типа 71%,
сибирского 15%, транспалеарктов 11%).
Подтипы
населения:
4.4 – сосновых,
темнохвойно-сосновых и берёзово-сосновых лесов и верховых болот от средней
тайги до лесостепи включительно (остромордая лягушка, серая жаба, обыкновенная
и средняя бурозубки, красная полёвка; 32752; 237/102; европейского типа 88%);
4.5 – берёзово-осиновых лесов,
полей-перелесков, полей и займищ в пределах подтаёжных лесов и лесостепи, остепнённых лугов и степей в пределах лесостепи (остромордая
лягушка, обыкновенная чесночница, серая жаба, обыкновенная и малая бурозубки;
26355; 241/103; европейского типа 90%).
Классы
населения:
4.5.1 –
берёзово-осиновых лесов (остромордая лягушка, серая жаба, сибирская лягушка,
обыкновенная и малая бурозубки; 19507; 198/103; европейского типа 75%,
сибирского 16%);
4.5.2 –
полей-перелесков и полей (остромордая лягушка, серая жаба, обыкновенная чесночница,
обыкновенная бурозубка, полевая мышь; 17159; 112/80; европейского типа 85%);
4.5.3 – остепнённых лугов и степей (остромордая лягушка,
обыкновенная чесночница, узкочерепная полевка, обыкновенная бурозубка, полевая
мышь; 19028; 107/66; европейского типа 96%);
4.5.4 – займищ
(остромордая лягушка, обыкновенная чесночница, малая бурозубка, полёвка-экономка,
мышь-малютка; 40579; 177/76; европейского типа 86%).
Подтипы
населения:
4.6 – внепойменных
осоково-вейниковых низинных болот, неостепнённых
лугов и пойм крупных рек от средней тайги до лесостепи включительно
(остромордая лягушка, серая жаба, сибирский углозуб, обыкновенная бурозубка, сибирская лягушка; 103829;
282/115; европейского типа 93%);
4.7 – лесостепных свалок (остромордая лягушка, полевой
воробей, узкочерепная полёвка, грач, коноплянка; 13480; 43/38; европейского
типа 68%, транспалеарктов 13%).
4.8 – степных займищ (зелёная жаба, остромордая лягушка,
обыкновенная чесночница, малая и обыкновенная бурозубки; 82260; 130/81;
европейского типа 62%, средиземноморского 31%);
5. Степной тип населения (степной зоны,
кроме займищ; остромордая лягушка, обыкновенная чесночница, зелёная жаба,
обыкновенная полёвка, обыкновенный емуранчик; 30762; 184/92; европейского типа
88%).
Подтипы
населения:
5.1 – степей и лугов
(остромордая лягушка, обыкновенная чесночница, зелёная жаба, обыкновенная
полевка, обыкновенный емуранчик; 41031; 169/47;
европейского типа 91%);
5.2 – полей (полевой жаворонок,
обыкновенная чесночница, серый хомячок, полевой воробей, грач; 5594; 64/57;
транспалеарктов 48%, европейского типа 27%, средиземноморского 15%);
5.3 – сосняков (зелёная жаба,
остромордая лягушка, обыкновенная полёвка, обыкновенная чесночница, тундряная
бурозубка; 4597; 63/43; европейского типа 56%, средиземноморского 30%).
II. Система водно-околоводных сообществ
А. Северная
надтиповая группировка
Типы
населения:
6 – арктических озёр и заливов (морянка, белохвостый песочник, белолобая казарка, кулик-воробей, круглоносый плавунчик; 250; 33/18; арктического типа 96%);
7 – арктических рек (чёрная казарка, турухтан,
белохвостый песочник, кулик-воробей, серебристая чайка; 65; 16/10; арктического
типа 91%);
8 – морских заливов в пределах субарктических тундр
и лесотундры (лапландский подорожник, жёлтая трясогузка, чечётка, турухтан,
кулик-воробей; 326; 68/33; арктического типа 60%, транспалеарктов 15%,
сибирского типа 13%);
9 – субарктических тундровых озёр и рек (белая
трясогузка, белохвостый песочник, морянка, жёлтая трясогузка, чернозобая
гагара; 167; 53/24; арктического типа 45%, транспалеарктов 29%, сибирского типа
19%).
10 – лесотундровых озёр и рек (чирок-свистунок,
полярная крачка, морянка, чернозобая гагара, фифи; 201; 42/18; транспалеарктов
36%, арктического типа 34%, сибирского 25%);
Б.
Срединная надтиповая группировка.
Типы
населения:
11 – северотаёжных озёр и рек (шилохвость, чирок-свистунок, турухтан, белая трясогузка, свиязь; 302; 67/29; транспалеарктов 42%, сибирского типа 22%, арктического 19%);
12 – озёр и рек средней и южной тайги (береговая ласточка, перевозчик, белая трясогузка, чирки - свистунок и трескунок; 245; 117/27; транспалеарктов 82%).
Подтипы:
12.1 – крупных и средних рек
(береговая ласточка, сизая чайка, белая трясогузка, речная крачка, серая
ворона; 208; 95/23; транспалеарктов 86%);
12.2 – озёр и малых рек
(перевозчик, чирок-свистунок, береговая ласточка, чирок-трескунок, белая трясогузка;
282; 94/27; транспалеарктов 79%).
13. Тип
населения подтаёжных и лесостепных озёр и рек (на сплавинах – остромордая и
сибирская лягушки, обыкновенная чесночница, малая бурозубка, мышь-малютка;
16322; 212/85; европейского типа 87%).
Подтипы
населения:
13.1 – рек, водохранилищ и
пресных озёр (остромордая и сибирская лягушки, чесночница, обыкновенная
бурозубка, мышь-малютка; 17216; 202/79; европейского типа 87%);
13.2 – соленых озер (малая
бурозубка, мышь-малютка, полёвки - экономка и водяная, обыкновенная бурозубка;
2005; 92/56; европейского типа 36%, транспалеарктов 29%,
средиземноморско-китайского типа 14% и тундро-лесостепных реликтов 13%).
14. Тип населения степных озёр и
рек (остромордая лягушка, зелёная жаба, обыкновенная чесночница,
жёлтая трясогузка, лысуха; 20976; 122/76; европейского типа 55%,
средиземноморского 29%, транспалеарктов 14%).
Подтипы
населения:
14.1 – пресных озёр и рек
(остромордая лягушка, зеленая жаба, обыкновенная чесночница, водяная полёвка,
обыкновенная бурозубка; 36210; 106/65; европейского типа 62%,
средиземноморского 34%);
14.2 – солёных озёр (озерная
чайка, круглоносый плавунчик, пеганка, шилоклювка, шилохвость; 497; 64/35;
транспалеарктов 57% арктического типа 13%);
КЛАССИФИФИКАЦИЯ НАСЕЛЕНИЯ ЖИВОТНЫХ
ПО ЭНЕРГЕТИЧЕСКИМ ПОКАЗАТЕЛЯМ
(КОЛИЧЕСТВУ ТРАНСФОРМИРУЕМОЙ ИМИ ЭНЕРГИИ)
(с
проникновением на незастроенную сушу от тундр до северной тайги
включительно)
А. Северная
надтиповая группировка
1. Арктический тундровый тип населения (подзоны
арктических тундр; лидеры по энергетике: сибирский лемминг, турухтан,
серебристая чайка, кулик-воробей, чернозобик; трансформируется – 31 тыс.ккал/км2.сут).
Подтипы
населения:
1.1 – тундр,
поселков, пойменных лугов, ивняков (сибирский лемминг, серебристая чайка,
турухтан, кулик-воробей, рогатый жаворонок; 26);
1.2 – болот (чернозобик,
кулик-воробей, чёрная казарка, круглоносый плавунчик, гага-гребенушка;
54);
1.3 – тампов
(турухтан, шилохвость, белолобая казарка, сибирский лемминг, гага-гребенушка; 26).
2. Субарктический тундровый тип
населения (тундр и болот подзон субарктических тундр и лесотундры,
посёлков южной тундры и лесотундры, бугристых северотаёжных
болот; сибирский лемминг, тундряная бурозубка,
полёвка-экономка, белая куропатка, краснозобый конёк;
37).
Б.
Срединная надтиповая группировка
3. Бореальный
тип населения (редколесий, лесов, вырубок, гарей, полей-перелесков, болот,
пойм крупных рек и свалок в пределах лесотундровых редколесий, лесной и лесостепной
зон, лесов степной зоны и северотаёжных посёлков;
полёвки - экономка и красная, обыкновенная бурозубка, водяная полёвка, средняя
бурозубка; 63).
3.1. Подтип населения
лесотундровых и северотаёжных лесов, редколесий и
посёлков (полёвки - красная и экономка, средняя бурозубка, тёмная полёвка, тундряная бурозубка; 34).
Классы
населения:
3.1.1 – лесотундровых редколесий и
редкостойных лесов (красная полёвка, тундряная бурозубка,
овсянка-крошка, средняя бурозубка, полёвка-экономка; 21),
3.1.2 – северотаёжных
редколесий и редкостойных лесов (полёвки - красная, тёмная и экономка,
бурозубки – средняя и обыкновенная; 32);
3.1.3 – прочих северотаёжных
лесов (полёвки - красная и экономка, средняя бурозубка, темная полёвка, тундряная бурозубка; 45),
3.1.4 – посёлков (серая ворона,
полёвки - экономка и водяная, белая трясогузка и домовый воробей; 44).
Подтипы
населения:
3.2 – северотаёжных внепойменных болот,
кроме бугристых (полёвки – экономка, водяная и красная, чернозобый дрозд и
овсянка-крошка; 20);
3.3 – темнохвойных и
темнохвойно-мелколиственных лесов, облесённых низинных болот средней и южной
тайги; южнотаёжных мелколиственных лесов,
полей-перелесков, зарастающих вырубок и гарей (обыкновенная бурозубка, полёвки
– экономка и красная, средняя бурозубка и тёмная полевка; 109).
Классы населения:
темнохвойных и
темнохвойно-мелколиственных лесов
и облесённых низинных болот
3.3.1 – средней тайги
(обыкновенная бурозубка, красная полёвка, средняя бурозубка, серая жаба,
полёвка-экономка; 83);
3.3.2 – южной тайги (полёвки –
экономка и красная, бурозубки – обыкновенная и средняя, тёмная пол–вка; 144);
3.3.3 – южнотаёжных
мелколиственных лесов, полей-перелесков, зарастающих гарей и вырубок
(обыкновенная бурозубка, полёвки – экономка, водяная, рыжая и красная; 79).
Подтипы
населения:
3.4 – среднетаёжных зарастающих
вырубок, гарей и полей-перелесков (полёвки – экономка, красная и водяная,
бурозубки - обыкновенная и средняя; 95);
3.5 – сосновых,
темнохвойно-сосновых и берёзово-сосновых лесов и верховых болот от средней
тайги до лесостепи и среднетаёжных мелколиственных лесов (красная полёвка,
обыкновенная и средняя бурозубки, полёвка-экономка и серая жаба; 42);
Классы населения:
3.5.1 - лесов и лесостепных рямов
(красная полёвка, обыкновенная бурозубка, полёвка-экономка, средняя бурозубка,
водяная полёвка; 51);
верховых
болот
3.5.2 – средней тайги (средняя
бурозубка, остромордая лягушка, серая жаба, обыкновенная бурозубка, полёвка-экономка;
16);
3.5.3 – южной тайги и подтаёжных
лесов (средняя бурозубка, красная полёвка, обыкновенная и малая бурозубки,
полёвка-экономка; 19).
Подтипы
населения:
3.6 – степных сосняков
(обыкновенная полёвка, тетерев, сорока, большая синица, лесная мышь; 27);
3.7 – подтаёжных облесённых
низинных болот (рыжая полёвка, обыкновенная бурозубка, полевая мышь, красная и
обыкновенная полёвки; 87);
3.8 – внепойменных
открытых низинных болот от средней тайги до подтаёжных лесов (остромордая
лягушка, тёмная полёвка, бурозубки – обыкновенная и малая, полёвка-экономка;
57);
3.9 – подтаёжных займищ (полёвки
– водяная и экономка, обыкновенный хомяк, малая бурозубка и остромордая
лягушка; 34);
3.10 – лесотундровых и северотаёжных пойм крупных рек (полёвка-экономка, обыкновенная
бурозубка, остромордая лягушка, водяная и тёмная полёвки; 51);
3.11 – пойм крупных рек от
средней тайги до лесостепи, подтаёжных и лесостепных мелколиственных лесов и
полей-перелесков, лесостепных низинных болот (полёвки – водяная и экономка,
обыкновенная бурозубка, остромордая лягушка и красная полёвка; 90).
Классы
населения:
3.11.1 – подтаёжных и лесостепных
внепойменных мелколиственных лесов и полей-перелесков (водяная полёвка,
обыкновенная бурозубка, полевая мышь, полёвки – красная и экономка; 73);
3.11.2 – пойменных лесов (полёвки –
водяная и экономка, обыкновенная бурозубка, остромордая лягушка и красная
полёвка; 112);
3.11.3 – пойменных лугов,
кустарников (полёвки – экономка и водяная, обыкновенная бурозубка, остромордая
лягушка и тёмная полёвка; 78);
3.11.4 – низинных болот, займищ и соров (полёвки – водяная и экономка, остромордая лягушка,
обыкновенная бурозубка и красная полёвка; 92).
4. Степной тип населения
(внепойменных лугов, полей и степей лесостепной и степной зон; грач,
узкочерепная полёвка, остромордая лягушка, водяная полёвка, полевая мышь; 49).
5. Рудеральный лесостепной тип
населения (свалок; грач, остромордая лягушка, обыкновенный хомяк,
полевой воробей, сорока; 189);
6. Синантропный таёжный тип
населения (средне- и южнотаёжных
посёлков; домовый воробей, полёвка-экономка, обыкновенная бурозубка, водяная и
красная полёвки; 69).
II. Система водно-околоводных сообществ
А. Северная
надтиповая группировка
7. Тип населения морских заливов и озёр от
арктической тундры до лесотундры (морянка, чернозобая гагара, серебристая
чайка, чирок-свистунок, белолобая казарка; 28).
Подтипы
населения:
7.1 – морских заливов (морянка, серебристая чайка,
турухтан, лапландских подорожник, малый веретенник; 18);
7.2 –озёр (чернозобая гагара, морянка, чирок-свистунок,
белолобая казарка, шилохвость; 41).
8. Тип населения рек от арктической тундры до
лесотундры (серебристая чайка, чёрная казарка, чернозобая гагара, средний
крохаль, белолобая казарка; 6);
Подтипы
населения:
крупных рек
8.1 – субарктической тундры (серебристая и сизая чайки,
свиязь, шилохвость, белая трясогузка; 2),
8.2 - лесотундры (сизая и серебристая чайки, полярная
крачка, свиязь, малая чайка; 0,3),
8.3 - средних и малых рек (серебристая чайка, чёрная
казарка, чернозобая гагара, средний крохаль, белолобая казарка; 8).
Б.
Срединная надтиповая группировка
9. Тип населения озёр и рек от северной тайги до
подтаёжных лесов, лесостепных и степных крупных и
средних рек (шилохвость, чирок-свистунок, сизая чайка, хохлатая чернеть,
кряква; 16).
Подтипы
населения:
9.1 - крупных рек (сизая и озёрная чайки, кряква, серая
ворона, шилохвость; 8);
9.2 - средних и малых рек и озёр (шилохвость,
чирок-свистунок, хохлатая чернеть, лысуха и кряква; 20).
10. Тип населения озёр и малых рек лесостепи, степи и
степных займищ (лысуха, кряква, чирок-трескунок, красноголовый нырок, водяная
полёвка; 132).
III. Система населения застроенной суши
11. Синантропный подтаёжно-степной
тип населения (посёлков от подтаёжных лесов до степи; домовый воробей,
сизый голубь, грач, полевой воробей, скворец; 112).
СОПОСТАВЛЕНИЕ КЛАССИФИКАЦИЙ
Классификации, составленные по показателям численности и количеству трансформируемой позвоночными энергии, в целом похожи, хотя первая из них состоит из двух зональных рядов (систем) населения суши и водно-околоводных сообществ, а вторая - из трех. В последней население суши делится на две системы незастроенной и застроенной части. При анализе по численности население посёлков имеет весьма сходный облик с сообществами окружающих ландшафтов. В какой-то мере это связано с некоторым недостатком данных по мелким млекопитающим. Зверьки, из-за невозможности отлова сходным методом в строениях, где преимущественно обитают синантропные виды, представлены в основном видами-посетителями, живущими постоянно в окрестных местообитаниях. Птицы-синантропы держатся в основном вне строений и хорошо учитываются. В результате население всех посёлков по численности входит в те же типы, что и их окружение, и поэтому не отражено в классификации. При анализе по энергетическим показателям, оценка значимости птиц резко возрастает, а мелких млекопитающих и особенно земноводных по сравнению с ними – уменьшается. В итоге, представления о населении посёлков приближаются к биоценотическим и специфика их сообществ отражается лучше, чем по численности.
Таким
образом, население посёлков от арктических тундр до лесотундры формируется
только из видов-посетителей и почти не имеет синантропной специфики. От таёжных
подзон до степи население селитебных территорий отнесено к отдельным типам. При
этом, начиная от подтаёжных лесов, из этих типов формируется самостоятельная
система сообществ незастроенной суши, представленная, правда, единственным
типом.
В
системе населения незастроенной суши, по классификации
составленной по энергетическим показателям, лесотундровый и лесной типы,
выделявшиеся по численности, объединяются в единый бореальный тип населения. В
этот же тип теперь входят сообщества степных сосновых боров, по численности относившиеся к степному типу. Подтип населения
сосновых, темнохвойно-сосновых, берёзово-сосновых лесов и верховых болот,
неделимый ранее, делится на сообщества лесов и лесостепных рямов, а также верховых
болот (отдельно средней тайги и южных подзон лесной зоны). Сообщества степных
займищ ранее входивших в лесной тип, более сходны с населением озёр и малых рек
лесостепи и степи за счёт сходства с сообществами озёрных сплавин и устьевых
займищ. Значительны различия в объёме степного типа. По энергетике в него
входят сообщества внепойменных лугов, степей и полей не только степной зоны,
как в классификации по численности, но и лесостепной, входившие в лесной тип
населения. Все эти отличия связаны, в основном, с большей значимостью птиц в
энергетическом балансе сообществ. Следует отметить также, что представление об
объёме некоторых типов и подтипов населения по энергетике стало более общим, т.е. имело меньшее число отклонений от обобщенных
представлений, чем по численности.
К
сожалению, в литературе есть лишь одна классификация, выполненная по населению
птиц и млекопитающих по территории, включающей Западную Сибирь [16]. Она
составлена по лесотундре, лесной и лесостепной зонам Сибири и Дальнего Востока.
По сравнению с нашей классификацией она не содержит данных по земноводным, но в
ней учтены сведения по крупным зверям (от бурундука и крупнее). Поэтому мы
сравниваем с ней нашу классификацию по энергетическим показателям, где значение
земноводных при разделении на таксоны менее существенно, чем по численности.
Принципы
этих классификаций существенно отличаются. Наша выполнена
по сходству населения на основании расчётов. При этом типами населения
считались группы вариантов первого разбиения. Примерно, по смыслу и территориально,
они совпадают с понятием типа растительности (лесной, луговой, степной), хотя
включают в себя и иные, но похожие варианты сообществ, например, лесной тип может включать население облесённых низинных болот, а
степной - полей, в том случае, если их сообщества сходны. Типы объединяются в
системы, если по максимальному или значимому сходству они следуют друг за
другом, образуя зональные ряды (например, на незастроенной суше тундровый тип
может сменяться лесотундровым, затем лесным и степным). Системы, как правило,
состоят из более похожих между собой типов и менее похожих на типы других систем,
например, незастроенной суши и водно-околоводных сообществ. В рядах по Западной
Сибири, как правило, прослеживается ослабление сходства между северными и более
южными типами. Это позволяет выделить внутри рядов надтиповые
группировки (группы типов).
Типы
тем же способом, что и вся совокупность доразбиваются
на подтипы, затем на классы, а иногда и подклассы. При всех
разделениях по сходству в объединившихся в группы вариантах ищется
фактор среды или их сочетание, одинаково проявляющийся на территории, где
проведены учёты животных. Эти факторы или режимы не должны также проявляться на
территории, занятой другой выделившейся группой. Например, совокупность по
сходству сообществ разделилась на три группы. В первую вошло население лесов, в
другую – открытых местообитаний, в третью – мозаичных, где лесные участки сочетаются
с открытыми, т.е. разбиение совпадает с изменением
облесённости. Это и является объяснением разбиения и основанием для названия
таксонов.
Следует
отметить, что сравнение нашей классификации и выполненной Н.Ф.Реймерсом
[16] из-за различий в принципах их составления, в объёме проанализированных
групп животных и площади территорий, к которым они относятся, некорректно. Оно
правомерно лишь в контексте: в какой степени обе
классификации учитывают дисперсию населения земноводных, птиц и мелких
млекопитающих, оценённую по энергии существования видов на территории Западной
Сибири в виде матрицы коэффициентов сходства. Поэтому суждения о том, какая из
классификаций лучше или хуже неуместны.
Н.Ф.Реймерс делил всю совокупность населения
птиц и млекопитающих вместе с растительностью. Сначала он экспертно
(умозрительно, не прибегая к расчётам) выделял элементарные фаунистические
комплексы геоботанических таксонов по сходству фауны, степени доминирования и
пребывания животных. Анализируя их сходство (тоже умозрительно), он пришёл к
выводу, что фаунистические комплексы, территориально относящиеся к одной
геоботанический формации более сходны между собой, чем с комплексами другой
формации. На этом основании автор выделяет зооформации
(население геоботанических формаций). Далее, тоже экспертно,
он убедился, что население групп, классов и типов растительных формаций внутри
себя тоже более похожи, чем на сообщества других формаций и т.д. Таким
образом, автор утверждает, что геоботаническое деление от растительной формации
и выше совпадает с фаунистическим (типолого-зоогеографическим).
Зооформации Н.Ф.Реймерс
делит по сходству на территориальные варианты, а их, в свою очередь, на
экологические варианты, например, по возрасту насаждений, и различает
образуемые вариантами природно- и антропогенно-сукцессионные
ряды. Итак, автор убеждён, что все границы таксонов геоботанической
классификации совпадают с границами животного населения, рассматриваемого
вместе с растительностью. Это фактически равнозначно навязыванию
геоботанических границ животному населению, поскольку степень их совпадения не
оценена количественно. На уровне формаций, как пишет Н.Ф.Реймерс,
контуры геоботанической и зоогеографической карт максимально приближаются друг
к другу, в то время как у нас это условно принято только на уровне наименьшей
единицы рассмотрения (ландшафтного урочища выделяемого типологически по
растительности). Таким образом, аксиоматически Н.Ф.Реймерс
считает границы растительной формации и животного населения совпадающими, хотя
и не везде, например, в лесостепи и поймах рек.
Наши
расчёты не подтверждают этого. Номинально тип зооформаций
Н.Ф. Реймерса близок к типу населения нашей
классификации, который, правда, нередко проникает на территорию, занимаемую
иным типом растительности, т.е. границы их не совпадают. Группа зооформаций и зооформация в
какой-то мере соответствует иногда рангу подтипов и классов населения в нашем
понимании, границы которых нередко совпадают с отличиями в составе
лесообразующих пород. Границы класса формаций на наших данных не выявляются.
Проявление территориальности вариантов формаций в нашей классификации
прослеживается в ранге надтиповых группировок. Таким образом, при некотором
сходстве в наборе факторов среды и их сочетаниях (природно-антропогенных
режимах) в рассматриваемых классификациях существенно различается иерархия их
значимости. Кроме того, классификация Н.Ф.Реймерса
внешне логически стройнее нашей, поскольку иерархия в ней больше соответствует
общности признака, а не силе и общности одновременно, как у нас. Наше
объяснение разделения на группы не совпадает с каким-то одним фактором или
признаком, например типом растительности, а включает, как правило, сочетание
факторов, иногда очень различных, но приводящих в
итоге к формированию сходных сообществ.
При
более подробном сравнении прежде всего обращает на
себя внимание отсутствие в классификации Н.Ф.Реймерса
понятия системы населения (незастроенной или застроенной суши и
водно-околоводных сообществ) и надтиповых группировок, т.е. соответствующие
типы населения (лесотундры, лесов, лесостепи, долин, водоёмов, посёлков и т.д.)
равнозначны и не объединены им в зональные ряды, которые лишь на уровне
территориального варианта делятся на северную и южную (срединную) части. Таким
образом, зональности на незастроенной суше, в посёлках и водно-околоводных
сообществах придаётся разный и меньший ранг, чем прослеживается в нашей
классификации.
В
тексте и на карте отображена лишь часть Западной Сибири - от лесотундры до
лесостепи включительно. Поэтому первым рассматривается лесотундровый тип населения,
в который входят сообщества тундр и лесов этой подзоны. В нашей классификации
имеется лишь лесотундровый подтип бореального типа, но в него входит население
лесотундровых и северотаежных лесов, редколесий и поселков. Сообщества
тундровых местообитаний относятся к субарктическому тундровому типу. Последнее
расхождение связано с тем, что на карте Н.Ф.Реймерса
тундры не отображены, хотя в тексте оговорены значительные отличия их населения
от остальной части лесотундры. Таким образом, фактическое отличие сводится к
рангу (тип-подтип) и объёму (включены или не включены сообщества лесов и
редколесий северной тайги и лесотундровых поселков).
Лесной тип населения по Н.Ф.Реймерсу ближе всего к бореальному в нашем понимании, хотя он меньше по объёму (не включает лесотундровых, лесостепных и долинных вариантов). Автор выделяет класс населения хвойных лесов, который не имеет аналогов в нашей классификации. Группы зооформаций темнохвойных лесов и их ближайших производных ближе всего к подтипу 3.3 нашего деления, хотя объём его иной. Так в нашей разбивке в него не входят сообщества северотаёжных лесов, но включено население облесённых низинных болот средней и южной тайги, южнотаёжных мелколиственных лесов и полей-перелесков. Эти сообщества отнесены Н.Ф. Реймерсом соответственно к болотному и лесостепному типу населения и группе зооформаций мелколиственных лесов. Группа зооформаций лиственничных лесов (вариант северотаёжных редкостойных), в нашем случае не выделяется в самостоятельный таксон, а входит в подтип лесотундровых и северотаёжных лесов, редколесий и посёлков (в класс северотаёжных). Группа зооформаций сосновых лесов делится на два варианта – северных и южных (в нашей интерпретации северной и срединной надтиповой группировок). Первый из них в нашем случае относится к «лесотундровому» подтипу бореального типа, а второй образует самостоятельный подтип вместе с населением верховых болот от средней тайги лесостепи и до среднетаёжных мелколиственных лесов. Группа зооформаций мелколиственных лесов единого таксона в нашей классификации не образует. Более богатые южнотаёжные сообщества ближе к населению темнохвойных лесов, а более бедные среднетаёжные – к сообществам сосновых лесов. Население подтаёжных и лесостепных мелколиственных лесов наиболее сходно с сообществами пойм крупных рек.
Лесостепной
тип населения Н.Ф.Реймерса разделился по нашей классификации
между рядом подтипов и классов, которые далеки по объёму от деления на варианты
зооформаций. То же следует сказать и про болотный тип
населения. Н.Ф.Реймерс пишет о необходимости и
одновременно невозможности деления сообществ болот по обводнённости, закустаренности и трофности и делит их только по зональной
принадлежности (лесотундровые и северотаёжные с одной
стороны, и более южные, с другой). Наше деление более дробное
и совпадает с различиями, связанными с трофностью, облесённостью и особенностями растительности. Меньшая
дробность классификации Н.Ф.Реймерса оговорена им и
связана с привязкой к карте масштаба 1:10000000.
Следующий
тип населения свойствен рекам и речным долинам (фактически рекам, мелким
озёрам, речным поймам и надпойменным лугам) и делится на северную
и южную части (лесотундрово-северотаёжную и более
южную). По нашей классификации такие сообщества суши входят в подтипы
бореального типа населения (3.10 и 3.11); объём второго подтипа больше, чем у Н.Ф.Реймерса. Он включает население подтаёжных и лесостепных
мелколиственных лесов и полей-перелесков, а также лесостепных низинных болот.
Сообщества лесостепных внепойменных лугов входят в
степной тип населения. Население рек и мелких водоёмов по Н.Ф.Реймерсу тоже делится на северную и южную часть, а в
нашей классификации относится к системе (ряду) водно-околоводных сообществ и
так же делится на северную и более южную части, хотя граница между ними проходит
южнее (северотаёжные варианты входят в срединную подтиповую группировку, а лесотундровые вместе с тундровыми
- в северную).
Население
побережий крупных озёр и морей Н.Ф.Реймерс выделяет в
отдельный тип населения. В нашей классификации эти сообщества тоже выделяются в
отдельный тип в пределах тундры и лесотундры, а более южные образуют единый тип
с сообществами рек и мелких водоёмов.
Отдельным
типом населения Н.Ф.Реймерс считает сообщества посёлков,
деля их на северные (лесотундры и северной тайги) и более южные (лесной зоны и
лесостепи). В нашей классификации прослеживается иное деление. Население
посёлков с севера до северной тайги включительно сходно с сообществами
окружающих ландшафтов и образует с ними единые типы. Средне-
и южнотаёжные посёлки образуют отдельный тип населения,
входящий в систему сообществ незастроенной суши и лишь от подтаёжных лесов до
степи включительно формируется специфическое населения ранга типа.
Таким образом, рассмотренные классификации существенно
отличаются, при этом умозрительная и в значительной степени гипотетическая классификация,
составленная Н.Ф.Реймерсом, менее дробна, в ней не
отражены взаимопроникновения разных типов и подтипов населения на нетипичную
для них территорию, например бореального (лесного) на облесённые болота и в
лесотундру или лесотундрового подтипа в пределы северной тайги. Однако в
них много общего. Информативность обоих классификаций в расчёте
на варианты сообществ Западно-Сибирской равнины (без тундровой и степной зон)
составляет 23 и 43 % учтённой дисперсии, т.е. соответствие имеющейся выборке
классификации Н.Ф.Реймерса почти вдвое меньше, чем
нашей. Следует отметить, что при расчётах использовано деление, не
только отображенное Н.Ф.Реймерсом в легенде карты и
тексте, но и, по возможности, дополнения, согласно приведённой автором
«таксономической системе эколого-зоогеографической типологии» (с. 19).
ПРОСТРАНСТВЕННО-ТИПОЛОГИЧЕСКАЯ СТРУКТУРА НАСЕЛЕНИЯ
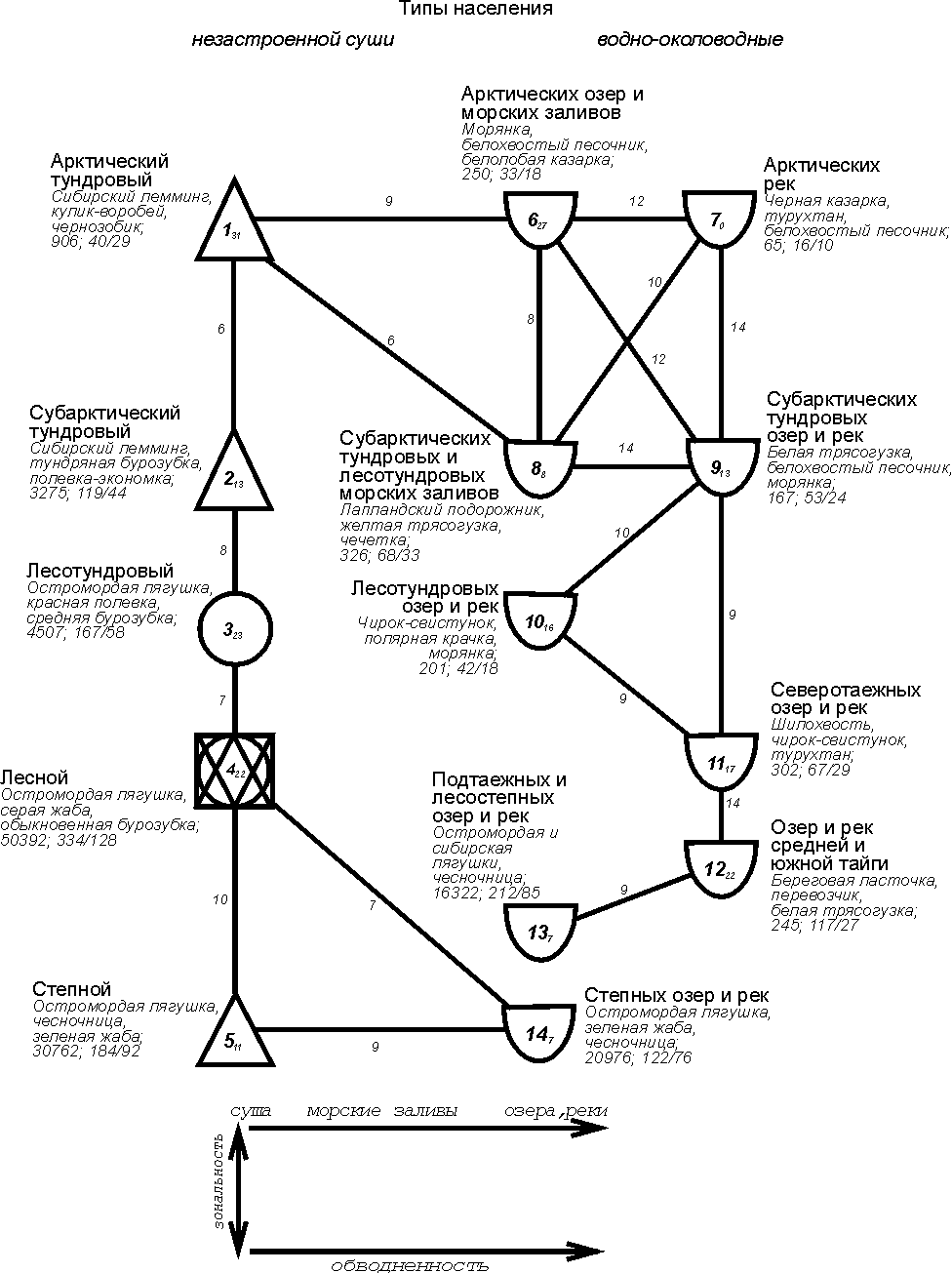
На
уровне типов населения по численности структура сравнительно проста и представлена
двумя зональными системами сообществ суши и водно-околоводных сообществ (рис.
1).
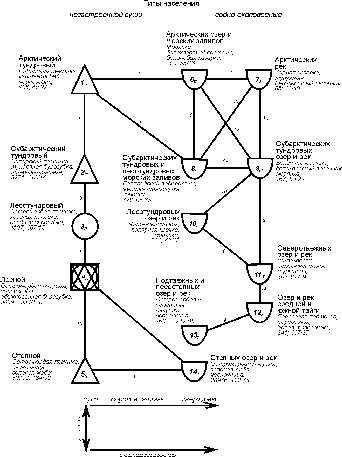
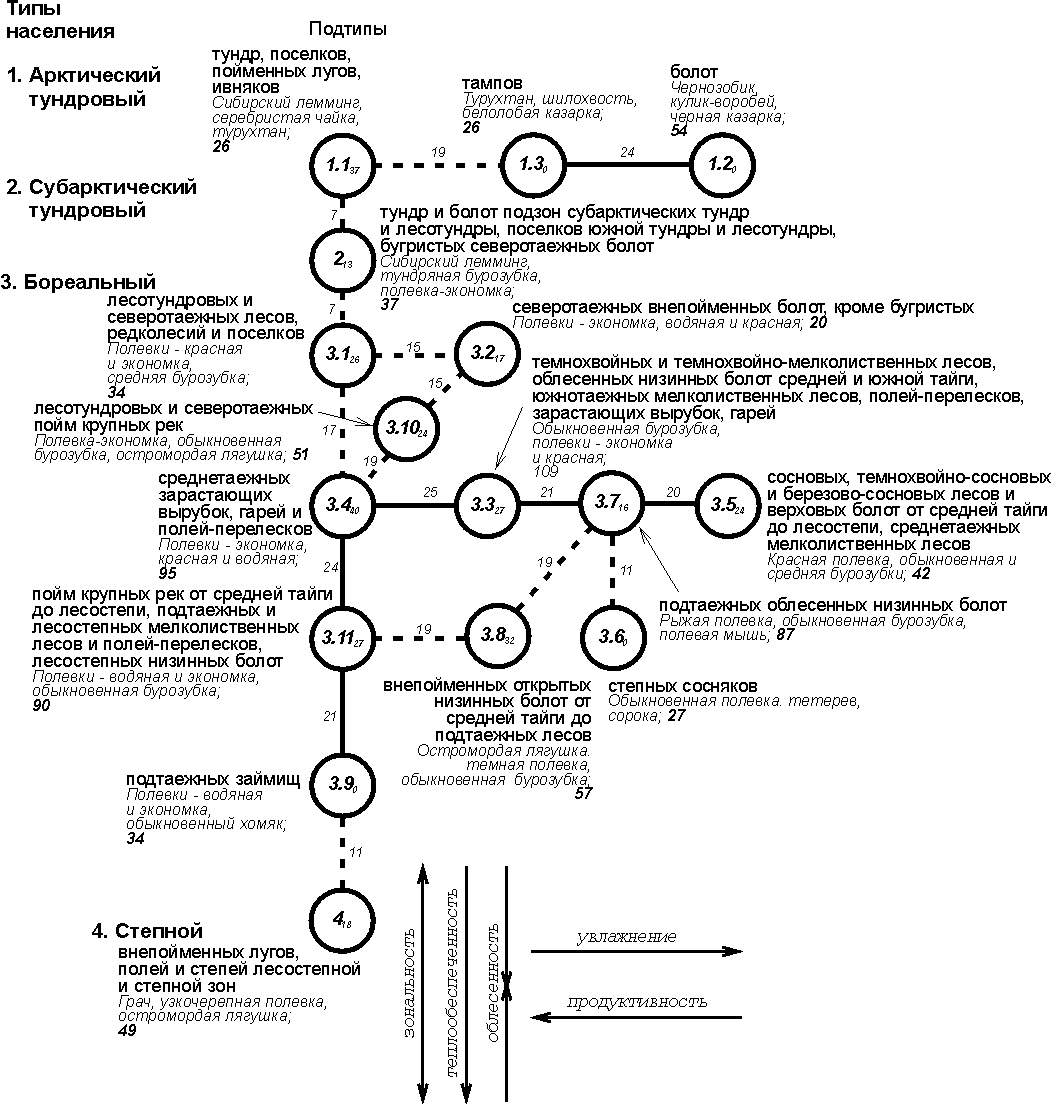
Рис. 1. Пространственно-типологическая структура населения
наземных позвоночных Западно-Сибирской равнины, 16 июля – 31 августа 1959-1997
гг. (по численности).
Индексами у номеров таксонов обозначено внутреннее сходство, у
связей между значениями – между ними. Запороговые
связи даны прерывистой линией. Рядом со значком таксона приведены три первых по
численности вида, плотность населения, число всех встреченных и фоновых видов.
На схемах 1 и 2 квадратами обозначены таксоны с преобладанием
сообществ лесов, кружками – тех местообитаний, где облесённые
участки чередуются с открытыми, треугольниками – открытых местообитаний
(основанием вниз – обедненных по продуктивности, а вверх – богатых), а полукольцом
- водных местообитаний.
Первая состоит из одного ряда, вторая - из
двух. Упорядоченность в обеих системах определяется зональностью, хотя и
существенно не совпадает с ландшафтными и геоботаническими представлениями о
ней. В системе водно-околоводных сообществ в северной части один из рядов
определяется отличием населения морских заливов от речного,
а в субарктической тундровой части и от озёрного. Начиная от лесотундры «двухрядность» графа, так же как и вертикальные изменения,
определяется зональностью. Обращает на себя внимание, что последний тип
населения степных озёр и рек, из-за значительно зарастания их, ближе к сообществам
суши, чем к своим более северным аналогам. Таким
образом, структура населения по численности определяется на уровне типов в основном
зональностью, обводнённостью и отличием водоёмов (морские или континентальные)
и водотоков.
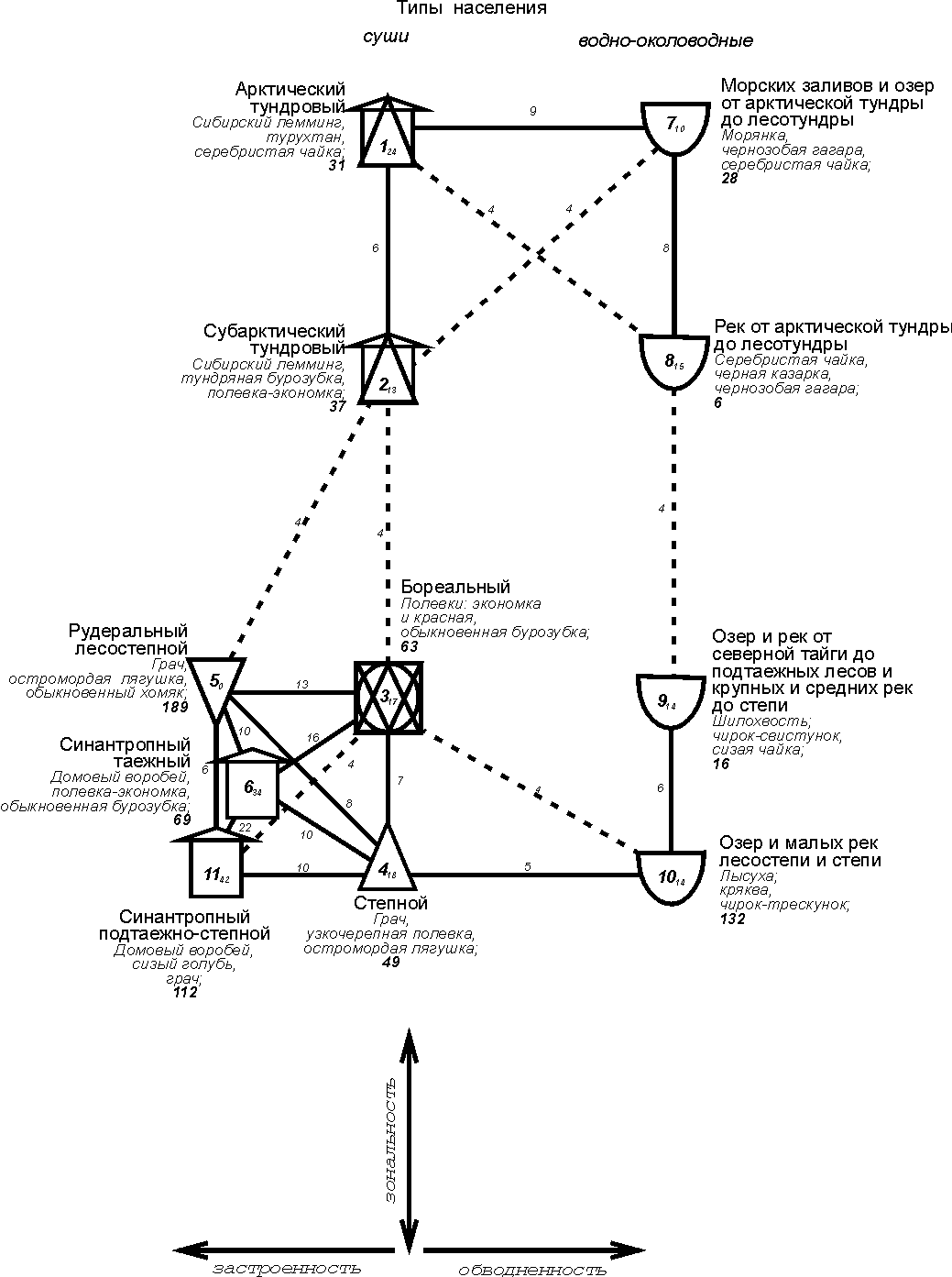
По
энергетике структура населения наземных позвоночных носит ещё более общий характер (рис. 2).
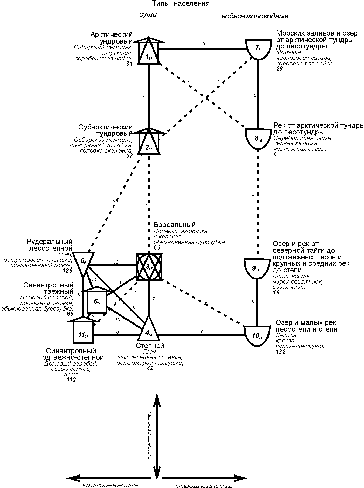
Рис. 2. Пространственно-типологическая структура населения наземных
позвоночных Западно-Сибирской равнины, 16 июля – 31 августа 1959-1997 гг. (по
количеству трансформированной энергии).
Условные обозначения как на рис. 1.
Граф состоит из двух параллельных зональных
рядов, в первый из которых входит население суши, во второй – водно-околоводные
сообщества. Первый ряд внизу графа имеет отклонение, связанное с большей
застроенностью южной части равнины. По сути, это ещё один ряд, усечённый до
одного типа. Севернее эти ряды сообществ суши смыкаются в один. Водно-околоводные
сообщества образуют один ряд, вместо двух как по численности, что связано с более общим характером классификации из-за увеличения энергетической
роли птиц. Отличия северной и срединной группировок в обеих системах по
энергетике больше, чем по численности – значимые связи между ними отсутствуют. На уровне подтипа отличия северной и срединной группировок и по
численности прослеживаются достаточно чётко.
В
водно-околоводных сообществах от арктических тундр до южной тайги включительно,
в связи с отсутствием сплавин, встречаются только птицы. Начиная от подтаёжных
лесов, за счёт высокой численности на сплавинах, лидируют земноводные. На суше в арктическом типе на первые места выходят мелкие
млекопитающие и птицы, в субарктическом тундровом типе – только первые, в
лесном – земноводные и мелкие зверьки, а в степном как ни странно – только
земноводные. Последнее связано с их высокой
численностью на степных болотах и поливных лугах и полях. По
энергетике в арктическом типе населения лидируют мелкие млекопитающие и птицы,
в субарктическом тундровом и бореальном только первые, а в степном –
представители всех исследованных групп позвоночных. В синантропном типе
населения и водно-околоводных сообществах лидируют исключительно птицы. На
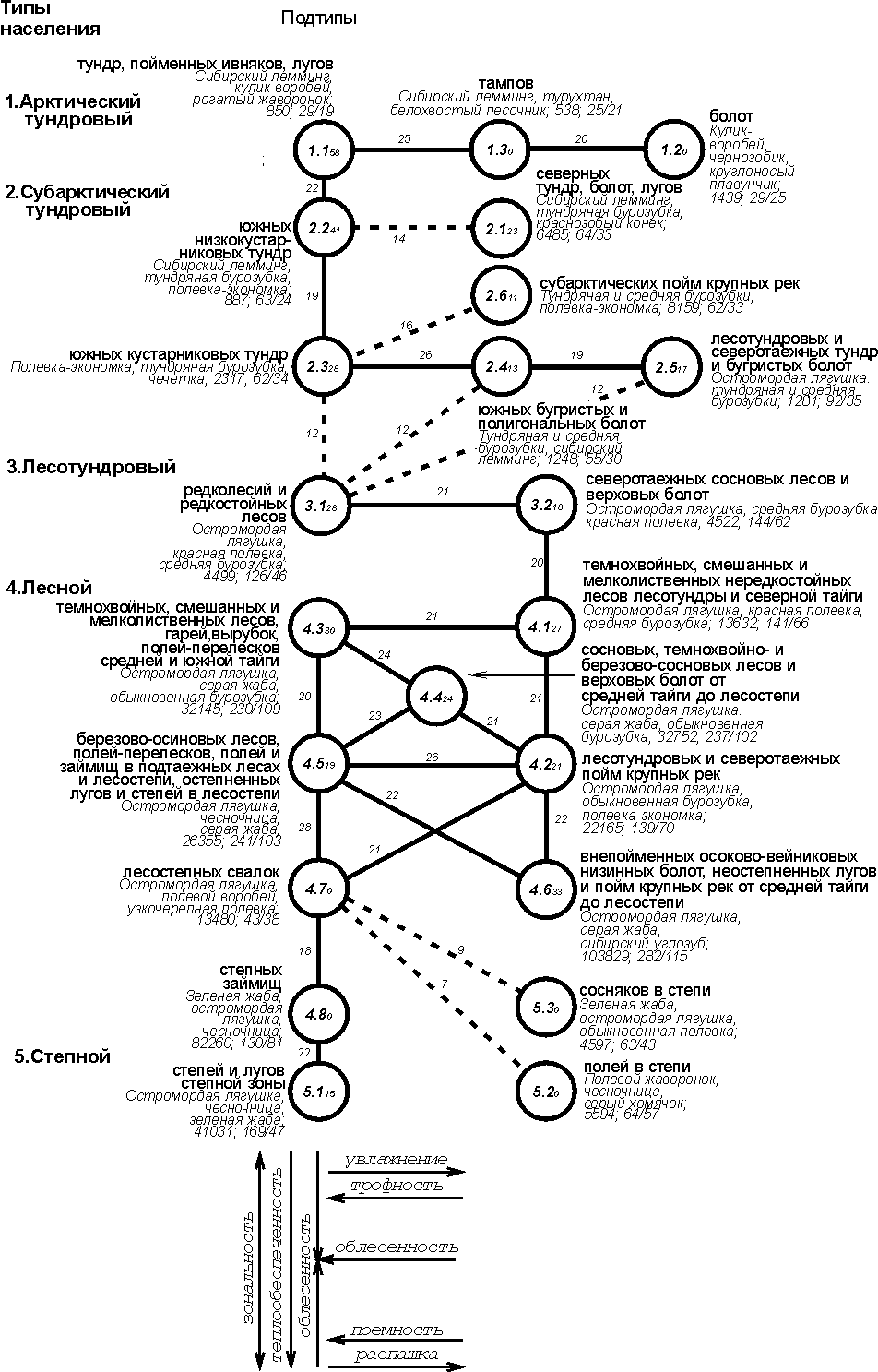
уровне подтипов по населению суши как по численности, так и по энергетике прослеживается в общем тот же характер изменений: зональные
(подзональные) отличия отображаются вертикальной
сменой, а внутризональные (внутриподзональные),
как правило – горизонтальной. На этих графах (рис. 3-4) отражено подтиповое деление сообществ только по ландшафтам
незастроенной суши.
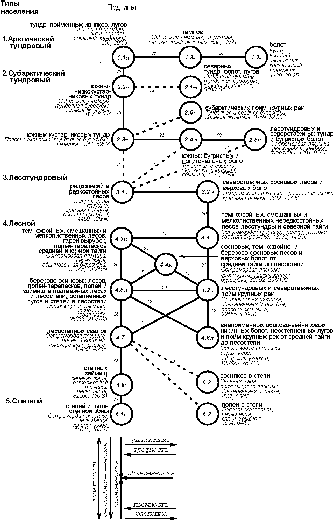
Рис. 3. Пространственно-типологическая структура населения
наземных позвоночных ландшафтов незастроенной суши Западно-Сибирской равнины,
16 июля - 31 августа 1959-1997 гг. (на уровне подтипа населения по
численности).
Условные обозначения как на рис. 1.
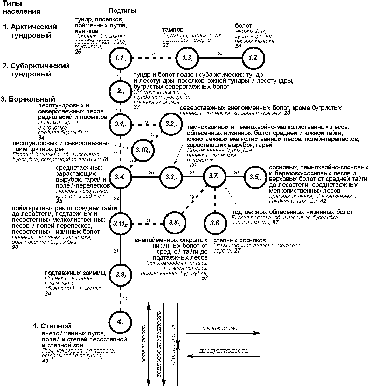
Рис. 4. Пространственно-типологическая структура населения
наземных позвоночных ландшафтов незастроенной суши Западно-Сибирской равнины,
16 июля – 31 августа 1959-1997 гг. (на уровне подтипа населения по количеству
трансформируемой энергии).
Условные обозначения как на рис. 1.
Структура
населения птиц по сравнению с таковой по всем позвоночным более
дробна [1]. По незастроенной суше, кроме тех же арктического и субарктического
тундровых и степного типов населения выделяются ещё типы: лесотундровый,
лесной, верхово-болотный, болотно-соровый и
лесостепных паров. По населению в целом все они, кроме паров, по энергетике
входят в бореальный тип, а сообщества лесостепных паров – в
степной. По численности, лесотундровый тип ещё сохраняет свою самостоятельность.
По застроенной суше в населении птиц прослеживается три типа населения, а по
энергетике - всего один. По численности синантропное население позвоночных не
образует типов. Количество типов водно-околоводных сообществ по птицам меньше,
чем для всех позвоночных как по численности, так и
энергетике.
По
мелким млекопитающим число типов в структуре населения[3]
равно таковому по численности всех позвоночных, а по энергетике превышает его
на один тип за счёт объединения лесотундрового и лесного типов населения.
Правда по мелким млекопитающим имеется лесостепной тип и единый тундровый тип
сообществ. По всем позвоночным последний делится на два самостоятельных типа
(арктический и субарктический), а лесостепной отсутствует вообще. По
численности всех позвоночных эти варианты населения входят в лесной тип, а по
энергетике сообщества облесенных пойменных и болотных ландшафтов этого типа
отходят к бореальному типу, а внепойменных лугов, степей и полей - к степному.
По мелким млекопитающим по структурному графу прослеживается влияние не только
зональности, но и облесённости, состава лесообразующих пород, увлажнения,
заливания в половодье и провинциальности. По всем группам позвоночных в сообществах
суши по численности проявляется влияние только зональности, а по энергетике ещё
и застроенности.
По
земноводным, по сравнению со всеми позвоночными, структура не только более дробна, но и сформирована совершенно иначе. Во-первых,
в ней чётче прослеживается провинциальность и особенности распределения доминирующих
видов – остромордой лягушки и серой жабы [5]. При этом лимитирование
по весьма различным факторам среды, может приводить к
высокому сходству сообществ в совершенно различных условиях по увлажнению,
теплообеспеченности и кормности. Это приводит к влиянию
зональности лишь в макроплане – сходное население
может сформироваться в одинаково влажных местообитаниях, но отличающихся по
кормности или теплообеспеченности и наоборот, различное в близких по кормности,
но различающихся по другим факторам. Это связано в основном с бедностью
видового состава земноводных, их высоким экологическим сходством и абсолютным
доминированием в населении 1-2 видов, в то время, как
другие группы позвоночных в видовом и экологическом отношении более разнородны,
чётче и постепеннее реагируют на весь комплекс ландшафтных условий, а не на
отдельные факторы среды.
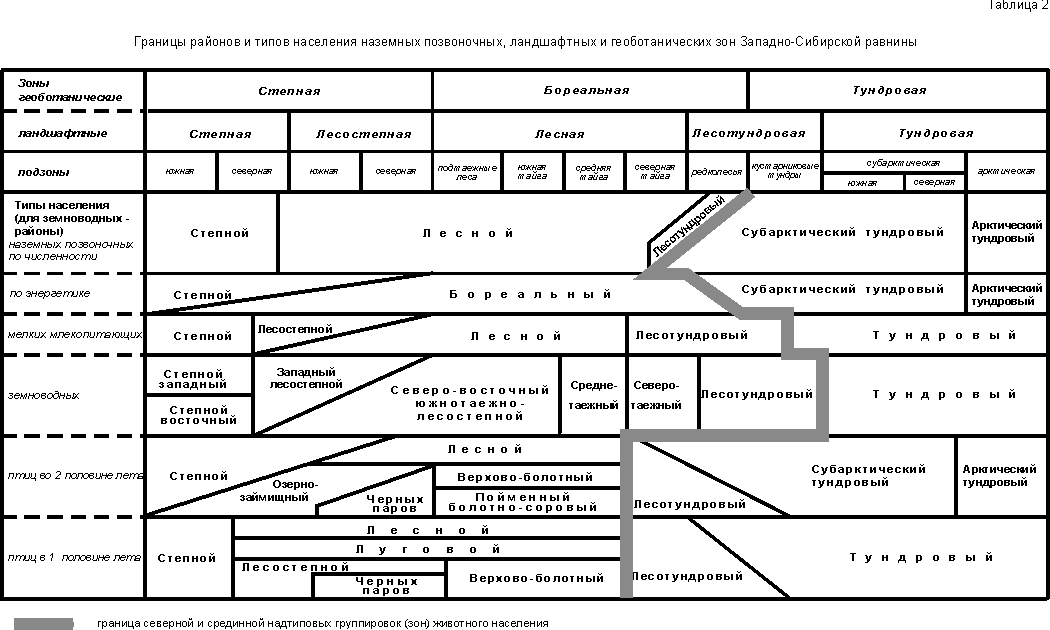
СОПОСТАВЛЕНИЕ ОСНОВНЫХ ГРАНИЦ НАСЕЛЕНИЯ ПОЗВОНОЧНЫХ С ЛАНДШАФТНЫМИ И ГЕОБОТАНИЧЕСКИМИ
Ландшафты
обычно подразделяют на три группы: незастроенной суши; селитебные и
промышленные; а также на водные (табл. 1). Растительность
тоже можно делить на водную, рудеральную, агроценозы
и естественную растительность суши, которая обычно делится на плакорно-зональную, болотную и пойменную.
{kind=link}
{kind=link}
{kind=link}
{kind=link}