Для мелких млекопитающих и земноводных
отклонение в значениях видового богатства в подтаежных лесах не выявлено. На
суходолах по всем изученным группам животных отдельно и по числу видов в сумме
широтные изменения идут также, как в целом по зонам (подзонам), а на болотах
прослеживается некоторое увеличение видового богатства птиц и мелких
млекопитающих в тундровой зоне.
В
поймах число отмеченных видов птиц и в целом трёх групп животных широтные
изменения идут также, а по мелким
млекопитающим минимум смещается в южнотаежные поймы, вероятно из-за интенсивных
половодий. Видовое богатство земноводных от лесостепи к тундрам уменьшается,
хотя и одинаково от северной тайги до тундр, а также в средней и южной тайге.
В
Северной Америке в целом неоднородность видового богатства млекопитающих выше,
чем на Западно-Сибирской равнине. В северной части материка изменения зависят
от широтной теплообеспеченности, а в южной - от высоты над уровнем моря,
ландшафтной неоднородности, удалённости от океана и влияния оледенения (Kerr, Packer, 1997).
В целом
по зонам (подзонам) в северной части Западно-Сибирской равнины (от тундр до
северной тайги включительно) большая часть населения изученных групп животных
приходится на мелких млекопитающих (58-84%), а южнее - на земноводных (68-89%).
В тундрах второе место по численности занимают птицы, участие земноводных
составляет менее одного процента. Второе место по видовому богатству в
лесотундре и северной тайге занимают земноводные, а начиная со средней тайги -
мелкие млекопитающие. В целом (с небольшими отклонениями) доля птиц и мелких
млекопитающих к югу убывает, а земноводных - увеличивается.
Доля
птиц в видовом составе животного населения составляет 83-92%, на мелких
млекопитающих приходится от 8 до 14%, а на земноводных - всего 1-3%. При этом
направленных изменений соотношения видов с запада на восток не прослеживается.
Итак,
по суммарным характеристикам животного населения в целом и его крупным
подразделениям имеются весьма четкие широтно-зональные градиенты (Чернов, 1975,
Чернов, Пенев, 1993). Однако, отдельным таксоценам и различным показателям в
совокупности свойственны индивидуальные отличия.
КЛАССИФИКАЦИЯ СООБЩЕСТВ
Птицы
В
классификации сообществ птиц Западно-Сибирской равнины прежде всего
прослеживаются отличия трех систем (рядов) населения: незастроенной или
застроенной суши и водно-околоводных местообитаний. При этом в первой половине
лета отличия этих рядов больше, чем во второй, когда ряды сообществ суши смыкаются
не только в пределах тундровой зоны, но и лесотундры, а открытые и зарастающие
озера и займища имеют сходное население. В каждом из рядов изменения сообществ
определяются, в основном, широтными отличиями в теплообеспеченности. Однако
границы выделяемых таксонов классификации сообществ не совпадают с ландшафтным
и геоботаническим делением территории (Карта «Растительность Западно-Сибирской
равнины», 1976; Чупахин, 1987). Так, ландшафтоведы делят Западно-Сибирскую
равнину на 5 зон, а геоботаники - на три. В сообществах же птиц чётко
прослеживаются две надтиповых группировки ранга зоны: северная и срединная.
Граница группировок по птицам проходит между северной и средней тайгой, т.е.
рассекает лесную (бореальную) зону. В северную группировку таким образом входят
орнитокомплексы тундровой и лесотундровой зон и северной тайги лесной зоны, а в
срединную - сообщества остальных подзон этой зоны, а также лесостепи и степи.
На застроенной территории эта граница несколько смещена к северу по крупным
северотаежным приречным старым поселкам, а на водоемах и водотоках она
смещается к северу на целую подзону, проходя между лесотундровыми редколесьями
и северотаежными редкостойными лесами. Смещение границ здесь связано с
отепляющим влиянием рек, текущих с юга. Надтиповые группировки делятся на типы
населения. При кластерном анализе они
выделяются при первом разбиении. Деление на типы наиболее информативно.
Надтиповые группировки и ряды выявляются по степени сходства типов населения.
Названия
типов сообществ и зональной растительности, как правило, совпадают, также как и
причины, определяющие их формирование. Но границы и объем типов населения, а
также ландшафтных зон не совпадают из-за автономности распределения животных и
растений и, до некоторой степени, различий в подходах к проведению границ:
типологический подход в разделении сообществ животных доминирует больше, чем
при классификации растительности и ландшафтов.
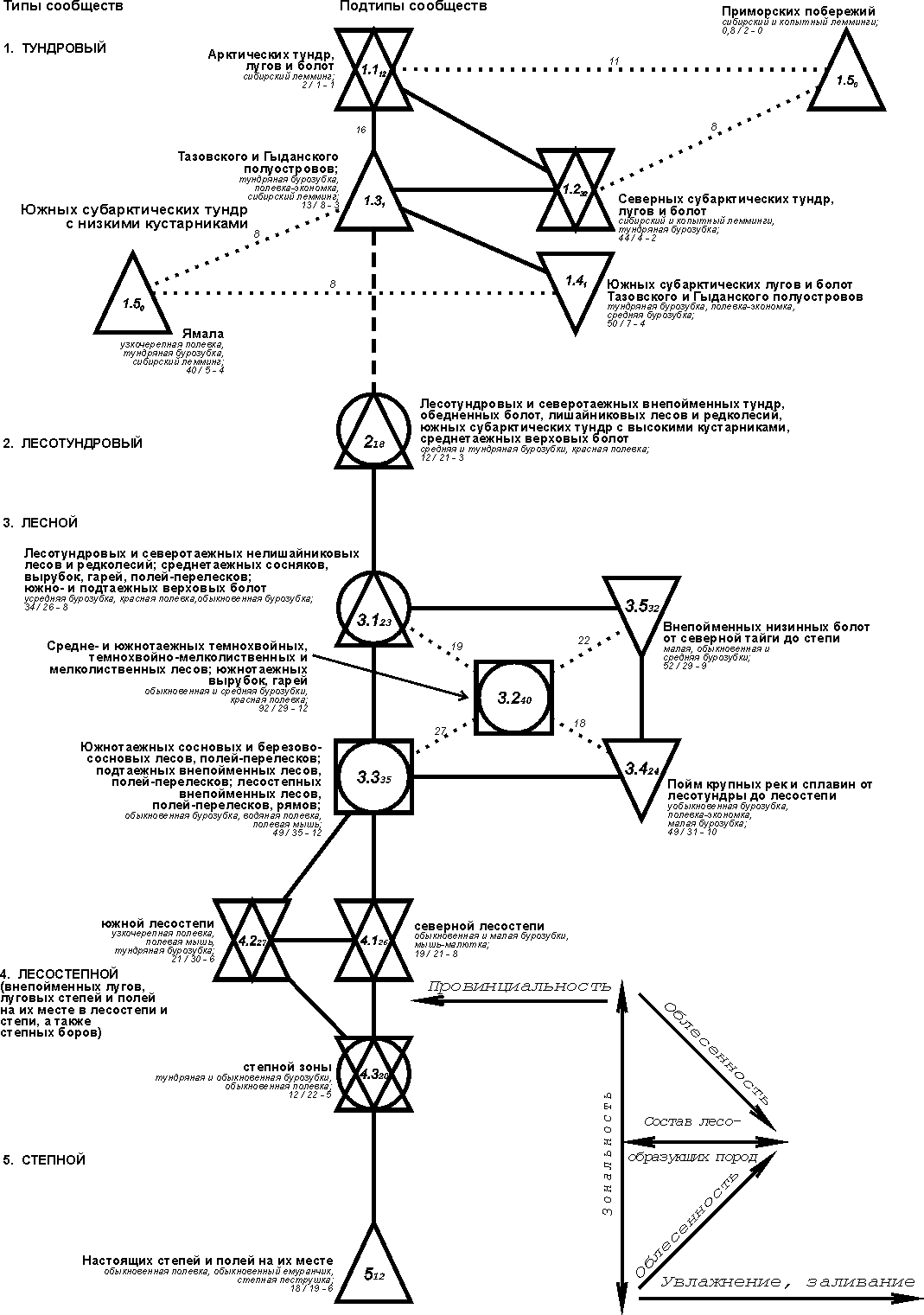
На
незастроенной суше во второй половине лета выделяется девять типов населения
птиц, на застроенной - три и в водно-околоводных вариантах - два. Северная
надтиповая группировка незастроенной суши состоит из двух тундровых типов
населения - арктического и субарктического, а также лесотундрового. Первый тип
населения занимает подзону арктических тундр, включая поселки; второй -
субарктические тундры (за исключением южных высоких кустарниковых) и проникает
в субарктические поселки, редкокустарниковые ерниковые и ивняково-ерниковые
кустарничково-лишайниково-моховые бугорковатые тундры в пределах лесотундровых
редколесий, а также на лесотундровые и северотаежные бугристые болота. Лесотундровый
тип населения включает сообщества полосы лесотундровых редколесий, кроме редко-
и низкокустарниковых тундр и бугристых болот. Он проникает в ерниковые тундры с
высокими кустарниками и ольхой, а также в ольховниковые южные субарктические
тундры и северную тайгу (кроме территории бугристых болот).
В
срединной группировке во второй половине лета выделяется 6 типов населения:
лесной, верхово-болотный, пойменный болотно-соровый, озерно-займищный, тип
сообществ лесостепных черных паров и
степной. Первый тип включает население лесов от средней тайги до степи, с
проникновением на зарастающие гари, вырубки, шелкопрядники, в перелески
чередующиеся с полями, а также пойменные луга-кустарники и на внепойменные
низинные болота (кроме лесостепных открытых низинных болот, полей с перелесками
южной лесостепи и степи). Верхово-болотный тип населения образуют сообщества
олиготрофных болот в пределах от средней тайги до лесостепи, а болотно-соровый
тип включает население пойменных открытых низинных болот и зарастающих пионерной
растительностью часто и надолго заливаемых в половодье соровых лугов в тех же
пределах. Озерно-займищный тип населения образуют сообщества займищ, озер и
заросшей части рек в пределах от подтаежных лесов до степи. Население
лесостепных черных паров из-за крайнего обеднения образует отдельный тип. В
степной тип входят орнитокомплексы лугов, степей, полей, выгонов лесостепной и
степной зон, а также полей, в том числе с перелесками и лесополосами южной
лесостепи и степи.
В
системе орнитокомплексов застроенной суши выделяются три типа населения -
лесотундровый, срединный и рудеральный. В первый входят сообщества
лесотундровых городов и поселков, а также северотаежных новых городов.
Срединный тип образуют варианты населения поселков и городов от средней тайги
до степи. К северу этот тип населения проникает в пределы северной тайги по
старым крупным приречным поселкам. Рудеральный тип включает орнитокомплексы
лесостепных свалок. В ряд водно-околоводных сообществ птиц входят два типа
населения - северный (тундрово-лесотундровый) и срединный (от северной тайги до
степи). Все типы делятся на подтипы, классы и нередко подклассы.
Классификация
населения, построенная по данным второй половины лета отличается меньшим числом
типов. В то же время, количество типов в ряду сообществ незастроенной суши на
один больше, поскольку единый тундровый тип населения разделен на два самостоятельных
- арктический и субарктический. В первой половине лета их ранг соответствовал
подтипу. При этом граница между ними проходила с небольшими отклонениями между
северными и южными субарктическими тундрами, а во второй половине лета - между
арктическими и северными субарктическими тундрами, т.е. севернее на одну
широтную полосу.
Это
связано с откочевкой из северных субарктических тундр ряда арктических видов
птиц к северу, а субарктических - к югу. Поэтому население арктических тундр
резче отличается от субарктических, в то время
как разница сообществ последних нивелируется. Дальнейшее деление в обеих
половинах лета сходно. Для второй половины лета характерна меньшая
дифференциация населения на уровне классов и подклассов из-за выравнивания
значений коэффициентов сходства, что обусловлено влиянием послегнездовых
кочевок.
В
лесотундровом типе населения во второй половине лета также прослеживается
меньшая дифференциация сообществ и некоторые отличия в иерархии значимости
отдельных природно-антропогенных режимов, но в целом изменения незначительны. В
частности, различия в орнитокомплексах пойменных лесов, лугов и болот сводятся
в первой половине лета к зонально-подзональным отличиям, а во второй - связаны
с увлажнением.
В
срединной надтиповой группировке изменения значительно больше. Так, лесной тип
населения распространяется на большую часть площади, занимаемой в первой
половине лета лесостепным и луговым типами. В лесной тип теперь входит
население не только облесенных низинных болот, но и закустаренных, слабооблесенных
и внепойменных открытых низинных болот и лугов. В качестве самостоятельного
типа населения от лугового остаются лишь сообщества пойменных открытых низинных
болот и соров. Причина в том, что лесные коньки (Antus trivialis) в значительном
количестве появляются в открытых и слабо облесенных низинных болотах и лугах,
хотя частично остаются в лесных и особенно в лесополевых местообитаниях.
Дубровники (Emberiza aureola), напротив, во второй половине лета чаще
встречаются в островных пойменных лесах, но часть их перемещается с лугов и
болот в поля с перелесками и на опушки лесов. Вместе с тем кочующие стаи синиц
(Parus) проникают в открытые местообитания по
кустарникам и лесным вкраплениям. Все это сближает население лесов и открытых
территорий, хотя чаще опосредованно, через сообщества промежуточных мозаичных
биотопов.
Орнитокомплексы,
составляющие в первой половине лета лесостепной тип, во второй половине делятся
между лесным и степным типами. В первый из них отходят орнитокомплексы полей с
перелесками в пределах подтаежных лесов и северной лесостепи, что обусловлено
распространением лесного конька и
дубровника. К степному типу населения относятся сообщества лесостепных лугов,
полей, луговых степей и полей-перелесков южной лесостепи, что связано с послегнездовым
увеличением численности грача (Corvus frugulegus) и полевого жаворонка (Alauda arvensis) после прикочёвки
их из подтаёжных лесов и северной лесостепи.
Входившие
в лесной тип орнитокомплексы лесостепных и подтаежных рямов (с болот, заросших
сосной), во второй половине лета включены нами в верхово-болотный тип, с
которым они в среднем наиболее сходны, хотя имеют значимое сходство как с
лесными, так и болотно-соровыми сообществами. Их промежуточное положение объясняется
тем, что большая часть лесостепных рямов представляет собой верховое болото, но
сосны выше, а насаждения гуще, чем на типичном низкорослом ряме. По периферии
таких местообитаний обычно тянутся узкой полосой более рослые сосновые рямы и
участки низинных кочковатых облесенных болот (согр). В рослых рямах и сограх
отмечается высокая численность птиц, что связано не только с большей, чем на
низкорослых рямах продуктивностью, но и с дефицитом лесов в лесостепи и подзоне
подтаежных лесов, большая часть которых сведена. За счет меньшей плотности
населения птиц на низкорослых рямах и более высокой - по периферии, в среднем в
орнитокомплексах значимую долю составляют лесные и лугово-болотные виды, что и
определяет наличие связей с соответствующими типами населения.
Кроме
того, в лесном типе населения становятся меньшими отличия сообществ степных
сосновых боров. В первой половине лета в лесном типе четче проявляются
подзональные отличия, во второй - пойменный режим и специфика западных лесов с
участием липы, а также отличия сообществ вырубок-гарей, полей-перелесков и облесенных
низинных болот от орнитокомплексов остальных лесных формаций.
Сообщества
займищных болот и заросших частей озер и дельт рек лесостепи и степи в первой
половине лета входили в качестве подтипа в луговой тип населения. Во второй
половине они образуют самостоятельный
озерно-займищный тип населения, в который входят орнитокомплексы займищных
болот, лесостепных и степных заросших тростником дельт и побережий рек, а также
озер, как заросших, так и открытых. Выделение этих орнитокомплексов в
самостоятельный тип связано с уменьшением доли камышевок (Acrocephalus) и
увеличением участия уток (Anas), что сближает
население займищ и открытых озер. С другой стороны, на займища не проникают
лесные коньки, кочующие синицы, дубровники, которые определяют возрастание сходства
слабо облесенных и закустаренных лугов и болот с орнитокомплексами лесов и
отделение их от займищных сообществ.
Во
второй половине лета возрастают различия между населением птиц застроенных и
рекреационных территорий. В водно-околоводных местообитаниях население птиц
средних по величине рек сближается с орнитокомплексами крупных рек, а
вовсе не малых, как это характерно для
первой половины лета.
Мелкие
млекопитающие
Классификация
населения мелких млекопитающих Западно-Сибирской равнины включает только одну
систему сообществ незастроенной суши. Ряд водно-околоводных вариантов
отсутствует, так как мелкие зверьки встречаются лишь на сплавинах озер, но
отмечены здесь те же, что и на суше виды. Население городов и поселков из
классификации исключено, т.к. зверьки не отлавливались в строениях. Сообщества
селитебных территорий вне строений представляют собой обедненные варианты
населения окрестных территорий с незначительным участием синантропов, поэтому
они входят в те же типы сообществ, что окружающие их населения. По
незастроенной суше, также как в населении птиц, выделяются две надтиповых
группировки сообществ - северная и срединная. Первая из них представлена одним
тундровым типом сообществ, вторая - четырьмя (лесотундровым, лесным,
лесостепным и степным). Границы выделенных типов также, как по птицам, не
совпадают с ландшафтными и геоботаническими. Распространение тундровой
растительности и сообществ северной надтиповой группировки мелких млекопитающих
и, соответственно, тундрового типа населения почти совпадают. Лишь часть
территориальных комплексов мелких млекопитающих южных субарктических
кустарниковых тундр (с высокими кустарниками) относятся к лесотундровому типу
срединной надтиповой группировки. Это, в какой-то мере отражает сходство с
ландшафтным делением, при котором часть южных субарктических кустарниковых
тундр вместе с полосой лесотундровых редколесий образуют самостоятельную
лесотундровую зону. При геоботаническом делении лесотундровые редколесья входят
в бореальную зону, а южные тундры - в тундровую.
По
населению мелких млекопитающих южная граница распространения северной
надтиповой группировки проходит существенно севернее, чем по птицам. Это
связано с тем, что ряд тундровых птиц в значительном количестве проникает в
лесотундру, а некоторые, свойственные последней, по обширным болотам - в северную
тайгу. В то же время, численность большей части настоящих таежников резко
снижается уже в северной тайге. Среди мелких млекопитающих сугубо тундровыми
можно считать только сибирского и копытного (Dicrostonyx torquatus) леммингов. Они
предпочитают суходольные ландшафты и по болотам к югу не продвигаются. Таежные
же виды мелких млекопитающих проникают в лесотундру и северную тайгу по
облесенным суходолам и даже в южные субарктические тундры с высокими кустарниками.
Поэтому их население ближе к лесному типу, чем к тундровому, и образует с
первым единую группировку. Входящий в нее лесотундровый тип населения
распространен в большинстве местообитаний лесотундровых редколесий и северной
тайги (кроме нелишайниковых лесов и редколесий, пойм и низинных болот) и
проникает к северу в южные субарктические тундры по участкам с высокими кустарниками,
а к югу - на среднетаежные верховые болота.
Лесной
тип населения занимает местообитания в основном от средней тайги до подтаежных
лесов, хотя проникает к северу по нелишайниковым лесам, низинным болотам и
поймам крупных рек вплоть до лесотундры и к югу по внепойменным лесам в пределы
лесостепи, а также на низинные болота, прибрежные озерные сплавины и поймы
крупных рек до степи включительно.
Лесостепной
тип населения распространен во внепойменных лугах, луговых степях и полях на месте
этих лугов и степей в пределах лесостепной и степной зон, а также в степных
сосновых борах. Степной тип свойствен настоящим степям и полям на их месте.
Бόльшая
часть типов населения представлена несколькими подтипами, а те, в свою очередь,
более, чем одним классом.
Земноводные
Классификация
сообществ земноводных также включает один ряд (систему) изменений населения, поскольку
только в южной части равнины на водоемах и реках встречается завезённая
случайно озерная лягушка, а сообщества поселков близки к населению окружающих
ландшафтов (синантропных видов среди земноводных нет). В населении амфибий
также как орнитокомплексах и сообществах мелких млекопитающих выделяются
северная и срединная надтиповые группировки. Первая из них занимает
арктическую, моховую и низкокустарниковую субарктические тундры, а также
тундры, бугристые болота и все неприречные светлохвойные леса и редколесья в
полосе южных кустарниковых субарктических тундр и лесотундровых редколесий.
Здесь земноводные нами не встречены, также как на песчаных побережьях и
островах соленого озера Чаны в лесостепи. Срединную группировку образует
население всей остальной территории, включая сообщества евтрофных и мезотрофных
болот и пойменных лугов южных субарктических тундр и лесотундровых редколесий.
Она делится на два типа. В первом доминирует серая жаба и/или остромордая лягушка,
во втором - зеленая жаба и/или обыкновенная чесночница. Второй тип занимает
западную часть подтаежных лесов, лесостепи и степи, а первый - всю остальную
часть территории равнины, где встречаются земноводные.
Граница
между северной и срединной группировками земноводных и мелких млекопитающих
почти совпадает, а по населению птиц проходит значительно южнее. Причины этого
явления различны. У земноводных они напрямую связаны с теплообеспеченностью, у
мелких млекопитающих и птиц определяются не столько с теплообеспеченностью,
сколько заболоченностью территории. Переувлажнение препятствует проникновению северных
видов мелких млекопитающих (сибирского и копытного леммингов) к югу по открытым
ландшафтам, у птиц, наоборот, способствуют смещению северных (гипоарктических)
видов к югу вплоть до северной тайги включительно.
Самое
дробное деление на типы и количество наименьших таксонов свойственно населению
птиц, меньшее - мелким млекопитающим и минимальное - земноводным. В
значительной степени это связано с числом видов в этих классах и их
экологическим разнообразием.
Зональные типы населения птиц формируются в значительной степени на основе предпочтения
видами благоприятных для них территорий. Тоже можно сказать про пойменный
болотно-соровый и займищный типы сообществ, а также о населении застроенной
суши и водно-околоводных местообитаний. Верховые болота и лесостепные черные
пары не предпочитаются ни одним из видов птиц. Их население состоит из видов,
свойственных иным местообитаниям, но изредка встречающихся в этих крайне бедных
ландшафтах на гнездовании, при случайных посещениях или залётах во время
кочёвок или за счёт проникновения птиц по вкраплениям пригодных для них
местообитаний. Все зональные типы сообществ мелких млекопитающих, кроме
лесостепного, тоже состоят из видов предпочитающих соответствующие территории.
Лишь последний формируется как за счет предпочтения, так и обеднения при
сочетании лесных и степных форм. Формирования обедненных вариантов за счёт
видов-посетителей не отмечено. Пространственную структуру населения земноводных
определяет убывание численности доминирующих видов.
ПРОСТРАНСТВЕННО-ТИПОЛОГИЧЕСКАЯ СТРУКТУРА
НАСЕЛЕНИЯ
Птицы
Для
сопоставимости с другими группами животных здесь мы рассматриваем структуру
основных изменений сообществ птиц только по второй половине лета на уровне
типов населения и лишь для незастроенной суши. Структурный ряд типов
орнитокомплексов чётко иллюстрирует зональное изменение сообществ: арктический
тундровый тип населения последовательно сменяется к югу субарктическим тундровым,
затем лесотундровым и лесным. Последний далее, правда опосредованно, переходит
в степной тип населения (рис. 1).
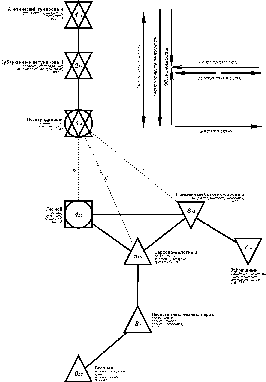
{kind=link}
Рис. 1. Пространственно-типологическая структура населения птиц незастроенной суши Западно-Сибирской равнины (на уровне типов). Вторая половина лета 1936-1995 гг.
Схемы 1-3 построены в примерном обратном масштабе: чем выше межклассовое сходство, тем ближе относительно друг друга расположены значки с номерами типов, подтипов или классов и индексами, обозначающими внутриклассовое сходство. Меж- и внутриклассовое сходство рассчитано как среднее от коэффициентов сходства соответствующих вариантов населения. Сила межклассового сходства показана лишь между первым и вторым классами и в случаях запорогового сходства. Если связь невозможно отразить в масштабе, она изображена пунктирной или штриховой линией и тоже имеет оценку сходства.
Квадратами обозначены классы с преобладанием сообществ лесов, кружками - тех местообитаний, где облесенные участки чередуются с открытыми, а треугольниками - открытых местообитаний (основанием вниз - с бедной продуктивностью, а вверх - с богатой). Рядом со значком класса приведены три первых по численности вида, плотность населения и число фоновых видов (по земноводным, на рис. 3, всех встреченных видов, их обилие и плотность населения).
Отклонения от ряда этих зональных типов сообществ связаны со значительными отличиями в продуктивности биоценозов как в сторону естественного увеличения (на внепойменных низинных болотах и в пойменном в болотно-соровом типе южной части равнины), так и уменьшения, - естественного на верховых болотах и антропогенного на черных парах. Таким образом, основные изменения облика населения определяются зональностью (теплообеспеченностью), увлажненностью, продуктивностью биоценозов и облесенностью местообитаний.Плотность
населения убывает от лесного типа к северу и югу, хотя в арктическом тундровом
типе суммарное обилие выше, чем в субарктическом. В отклонениях от основного
ряда плотность населения увеличивается с ростом продуктивности биоценозов и
уменьшается с обеднением ее.
Количество
фоновых видов изменяется почти также как и плотность населения, но без
отклонения в арктическом типе и без увеличения в болотно-соровом. Последнее
связано с меньшим разнообразием болот и соров по сравнению с набором
местообитаний лесного типа.
Мелкие
млекопитающие
Граф
сходства населения мелких млекопитающих
составлен на уровне подтипа, поскольку меньшее число подтипов, чем по птицам,
позволяет построить достаточно иллюстративную и в тоже время простую схему (рис. 2).
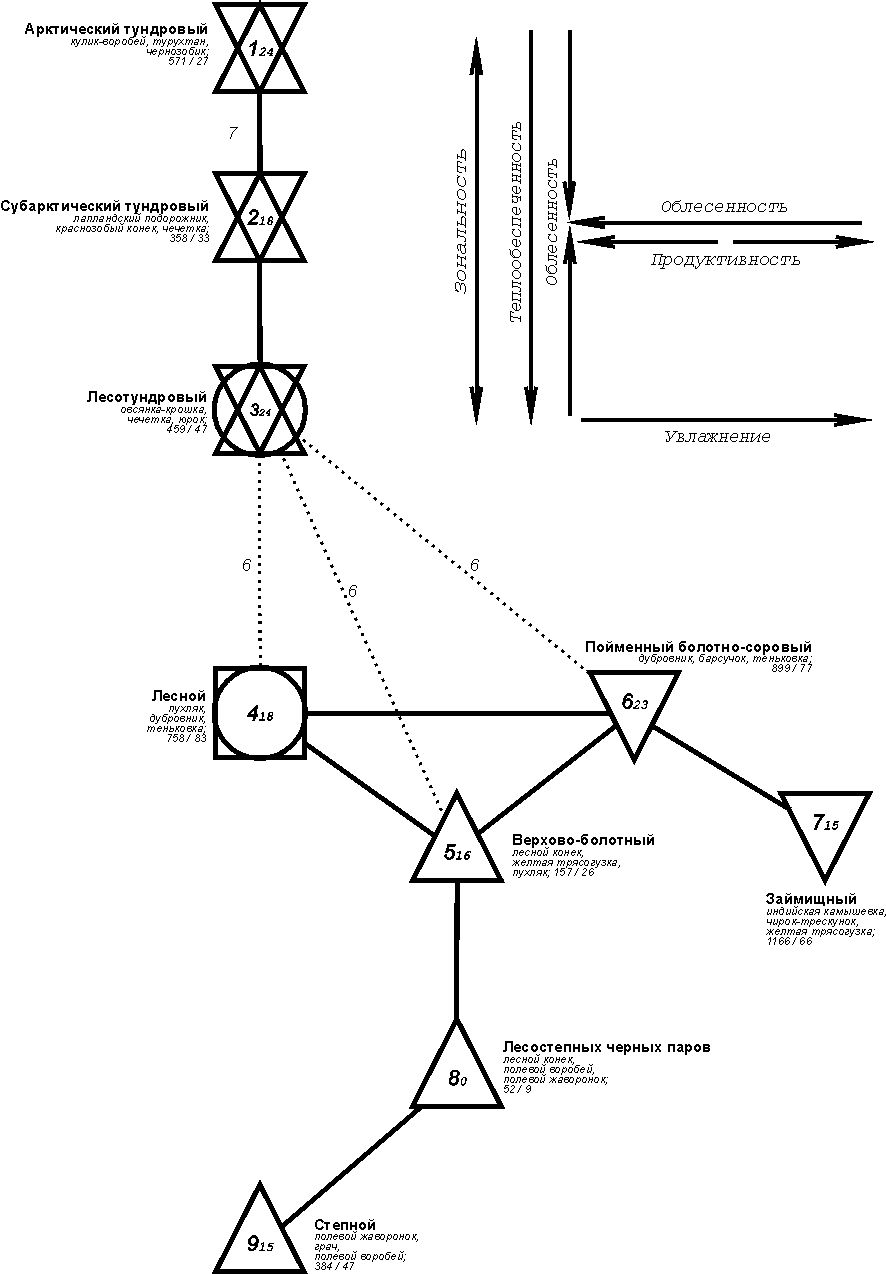
{kind=link}
Рис. 2. Пространственно-типологическая структура населения мелких млекопитающих Западно-Сибирской равнины (на уровне подтипа). Вторая половина лета 1965-1995 гг.
Условные обозначения как на рис. 1.
На схеме четко прослеживаются тренды населения, связанные с широтным изменением теплообеспеченности (зональностью), увлажнением, облесенностью, составом лесообразующих пород и слабо компенсируемым половодьем. Не менее чётко видны сгущения подтипов, относящихся к одному типу, а также отсутствие значимой связи между северной и срединной группировками (соответственно типы 1 и 2-5). Правда, связь подтипов 1.5 и 1.6 с остальными из этого типа еще меньше, но это или крайний вариант обеднения сообществ, связанный с периодическим нагонно-приливным затоплением побережий морских заливов, или локальные сообщества Ямала с преобладанием узкочерепной полевки (Microtus gregalis).В целом
плотность населения, видовое богатство и число фоновых видов наиболее высоки в лесном типе населения и убывают к северу
и югу, хотя по суммарному обилию эта тенденция несколько нарушается в южных
субарктических тундрах, где, как уже говорилось, в один из трех полевых сезонов
отмечено массовое размножение сибирского лемминга. Некоторое уменьшение
плотности населения прослеживается и при переувлажнении в пределах лесной зоны,
а также при распашке и при влиянии половодья, кроме того в светлохвойных и
лишайниковых лесах зверьков меньше, чем в
нелишайниковых, темнохвойных, смешанных и мелколиственных.
Таким
образом, общее направление изменений населения мелких млекопитающих и птиц
сходно и определяется, в основном, теплообеспеченностью (зональностью). Однако
отклонения в зонально-подзональных рядах, связанные с отличиями в
продуктивности для населения птиц проявляются в ранге типа сообществ, а для
мелких млекопитающих – подтипа. Во время послегнездовых кочевок бόльшие миграционные способности птиц
приводят к доминированию населения лесного типа на большей территории не только
в лесных ландшафтах, но и в открытых и частично облесенных местообитаниях
лесной зоны и лесостепи. У зверьков реакция на затопление чётче, чем у птиц.
Проникновение сообществ мелких млекопитающих лесного типа к югу в южной
лесостепи и степной зоне на территорию переувлажненных ландшафтов больше, чем
лесных и полуоблесенных. Кроме того, в суходольных ландшафтах
макрооблесенность, т.е. общий характер облесения, на зверьках сказывается
больше, чем на птицах. Это также связано с большей способностью последних к перемещению.
Земноводные
Пространственно-типологическая
структура населения амфибий лишь в самых общих чертах, в ранге надтиповых
группировок иллюстрирует связь изменений с зональностью (земноводные обитают в
срединной и южной частях равнины, на севере их почти нет). На графе
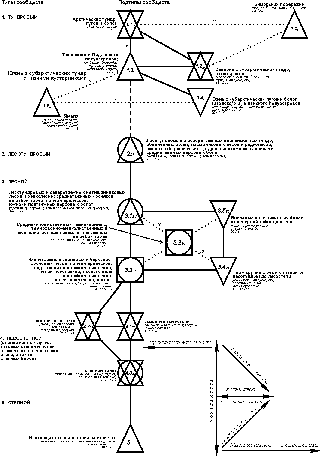
прослеживаются три узла с максимальной плотностью населения (рис. 3).
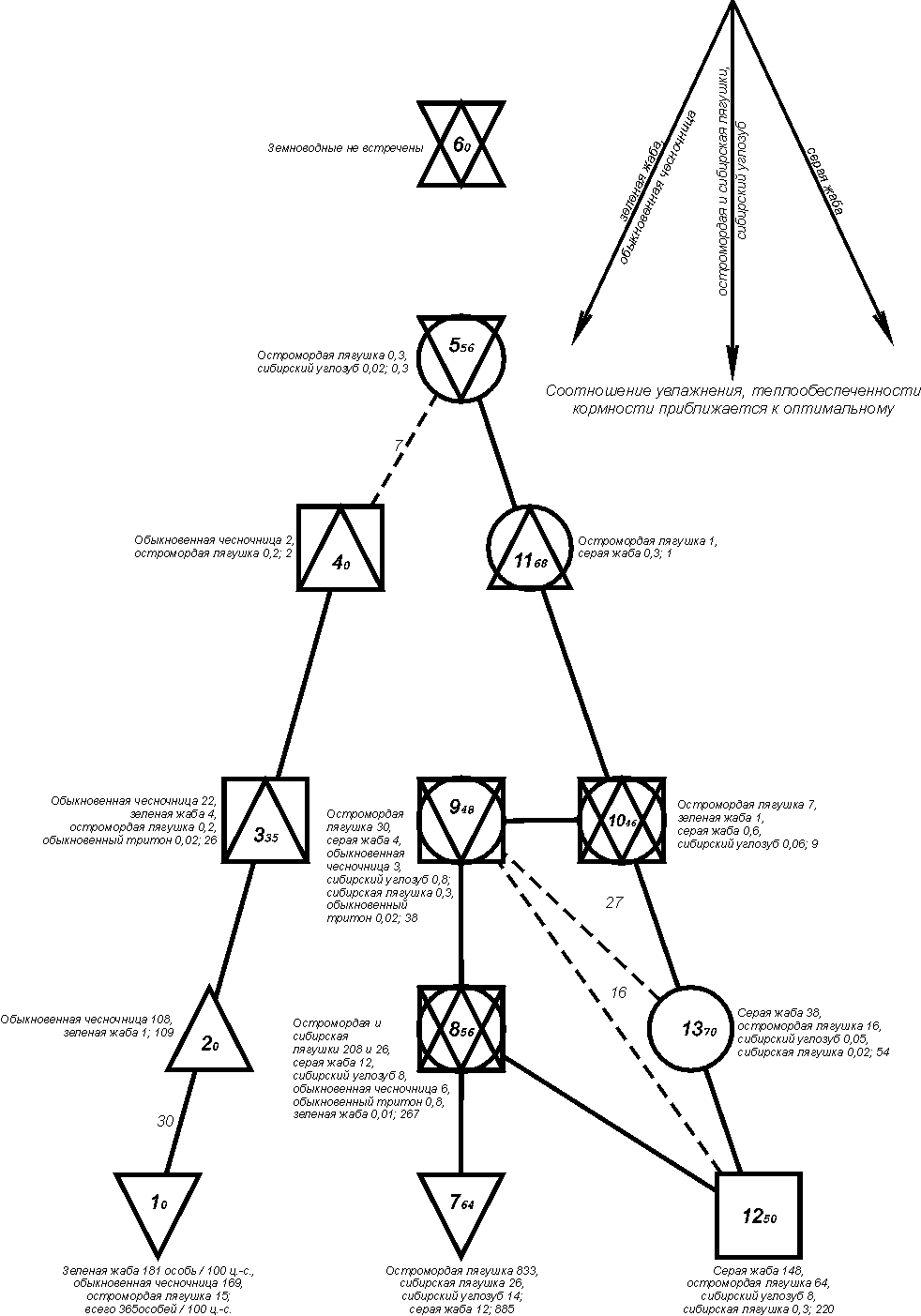{kind=link}
Рис. 3. Пространственно-типологическая структура населения земноводных Западно-Сибирской равнины (на уровне класса населения).
Условные обозначения как на рис. 1.
Варианты населения по классам: 1 - влажных лугов, займищ и полузаброшенных поселков среди них в южной части степи; 2 - разнотравно-полынных степей средней и южной части степной зоны; 3 - сосняков, светлохвойно-мелколиственных лесов, посадок, рямов западной части лесостепи; типчаково-полынных и песчано-ковыльных степей средней и южной части степной зоны; 4 - сосняков и полей средней части степной зоны; 5 - лишайниково-осоково-моховых тундр и болот южной кустарниковой субарктической тундры; лесотундровых редколесий и редкостойных лесов долины реки Таз; внепойменных суходольных местообитаний северной тайги; 6 - территорий, не населенных земноводными (арктическая, моховая и низкокустарниковая субарктическая тундра; тундры, бугристые болота и все неприречные светлохвойные леса и редколесья лесотундровой зоны; песчаные побережья и острова соленого озера Чаны в лесостепи); 7 - соров и низинных болот поймы Оби и Иртыша в южной и средней тайге; 8 - низинных болот поймы Оби и Енисея в северной тайге и подтаежных лесах; лугов, ивняков, осинников поймы Оби и Иртыша в средней тайге и подтаежных лесах; внепойменных низинных открытых болот южной тайги и подтаежных лесов; подтаежных облесенных низинных болот и пойменных полей-перелесков; подтаежных заболоченных (хотя бы частично) лесов долины Тобола; лесостепных влажных: лесов, лугов, луговых степей, поливных полей многолетних трав и евтрофных болот, пойм крупных рек; лугов, займищ, сплавин озер и прибрежных тростниковых зарослей в северной и центральной степи; 9 - лесотундровых лесов поймы р.Таз; лугов, ивняков и соров поймы Оби и Енисея в северной тайге; среднетаежных внепойменных темнохвойных и мелколиственных лесов и их производных; внепойменных низинных и переходных болот и пойм притоков крупных рек и Енисея в средней тайге; лугов, ивняков, осинников поймы Енисея, Оби и Иртыша в южной тайге; внепойменных суходольных местообитаний южной тайги, подтаежных лесов и лесостепи (кроме лесостепных поливных полей многолетних трав, а также сосновых и березовых лесов западной части подзоны); подтаежных рямов; центральной и северной части степи (кроме полей, чередующихся с перелесками); 10 - осоковых болот южной субарктической тундры; лесотундровых комплексных болот и поймы реки Ныда; северотаежных приречных влажных и полузаболоченных (несветлохвойных) лесов, редколесий и внепойменных болот северной тайги; сосняков и светлохвойно-мелколиственных лесов восточной части лесостепи; сосняков южной части степной зоны; 11 - поймы Оби (кроме тундр), тампов, приречных редкостойных лесов и редколесий в лесотундровой зоне; южнотаежных верховых болот; 12 - сосняков, их производных и рослых рямов средней тайги; внепойменных заболоченных облесенных неолиготрофных местообитаний южной тайги; полей-перелесков центральной и восточной части северной степи; 13 - среднетаежных верховых болот; лесостепных березово-осиновых лесов, коллективных садов и полей-перелесков на месте сосновых приобских боров.
В первом из них (класс 1) численно преобладают зеленая жаба и обыкновенная чесночница (50 и 44%), во втором - абсолютно доминирует остромордая лягушка (класс 7; 94%); в третьем - серая жаба (класс 12; 87%). От этих центров идет уменьшение обилия перечисленных видов земноводных, что является следствием снижения теплообеспеченности, продуктивности биоценозов и увлажнения, а для зеленой жабы и обыкновенной чесночницы - удаления от оптимума ареала и уменьшения к северу степени аридизации. У обыкновенной чесночницы, кроме того, отмечена тенденция уменьшения обилия с запада на восток. На численность серой жабы и сибирского углозуба и в меньшей степени лягушек отрицательно сказывается половодье. Лягушек в поймах на единицу площади больше, чем на внепойменных суходолах. Здесь выше влажность и больше пригодных для выплода водоемов. Тем не менее, на некоторых незаливаемых низинных открытых болотах лягушек больше, чем на заливаемых.Снижение
обилия зеленой жабы и обыкновенной чесночницы идет в ряду классов 1-6;
остромордой и сибирской лягушек, а также сибирского углозуба - в классах 7-11 и 5; серой жабы - в ряду 12, 13, 10, 11,
5, 6. Все ряды на графе сходятся к 6-ому классу, составленному пробами, где
земноводные не встречены.
Как
видно по составу классов весьма различные факторы приводят к формированию у
земноводных сходного населения. Так же как в большинстве зон и подзон, где
население амфибий рассмотрено отдельно, структура сообществ совпадает с
уменьшением обилия доминирующих видов. Эмерджентность и целостность населения
земноводных невелика из-за невысокого видового разнообразия. Пространственно-типологическую
структуру их населения определяет, в основном, сочетание теплообеспеченности,
увлажнения и кормности для четырех доминирующих видов. Кроме того, прослеживается
как положительное, так и отрицательное прямое и косвенное влияние половодья.
ПРОСТРАНСТВЕННО-ТИПОЛОГИЧЕСКАЯ ОРГАНИЗАЦИЯ
НАСЕЛЕНИЯ
При
кластерном анализе данных по населению исследуемых групп животных, составлении
классификаций и построении графов выявлен набор основных структурообразующих
факторов среды. Их список приведён в таблице 3.
Связи этих факторов с неоднородностью населения следующие. Для всех трёх групп
животных наиболее значима связь с зональностью (подзональностью), несколько меньше
- с облесенностью и составом лесообразующих пород. Сила и общность связи
территориальной изменчивости сообществ с остальными факторами среды, в том
числе и с антропогенным воздействием, существенно меньше. Последнее имеет
высокое локальное влияние, но в целом по Западно-Сибирской равнине мало
значимо.
Таблица 3.
Оценка силы и общности связи факторов среды и населения птиц, мелких млекопитающих и земноводных Западно-Сибирской равнины (вторая половина лета 1963-1995 гг.)
Фактор, режим
|
Учтенная дисперсия по неусредненным данным (средняя по восьмикратной выборке в 100 вариантов), % |
||
|
птицы |
мелкие млекопитающие |
земноводные |
|
|
Зональность (подзональность) |
41±0.5 |
31±1 |
15±0.7 |
|
Облесенность |
27±2 |
22±2 |
4±0.6 |
|
Состав лесообразующих пород |
16±1 |
19±1 |
5±0.6 |
|
Увлажнение |
7±0.7 |
3±0.7 |
0.9±0.2 |
|
Продуктивность |
4±0.3 |
4±0.4 |
2±0.2 |
|
Провинциальность |
3±0.8 |
5±0.5 |
1±0.2 |
|
Мезорельеф |
3±0.5 |
1±0.2 |
2±0.3 |
|
Антропогенное влияние, в том числе: |
2±0.3 |
2±0.4 |
2±0.3 |
|
застроенность |
1±0.3 |
0.4±0.3 |
1±0.3 |
|
Распашка |
0.5±0.2 |
1±0.08 |
1±0.2 |
|
Всего факторы |
62±1 |
45±2 |
23±0.5 |
|
Всего режимы |
61±1 |
27±1 |
13±0.5 |
|
Всего факторы и режимы |
69±0.6 |
47±2 |
24±0.5 |
Наиболее
тесная связь с основными структурообразующими факторами среды прослеживается
при анализе населения птиц (в целом учитывается 62% дисперсии). Показатели по
мелким млекопитающим почти в полтора раза меньше. Это уменьшение связано, в основном,
с большими, чем у птиц, годовыми колебаниями численности мелких млекопитающих.
Еще меньше сила и общность связи факторов среды и неоднородности населения
земноводных (почти втрое по отношению к сообществам птиц и вдвое - мелких
млекопитающих). Снижение учитываемой дисперсии определяется такими лимитирующими
факторами, как наличие водоемов выплода, локальных и соседствующих
переувлажненных мест, где земноводные могут пережидать засушливые периоды между
дождями или восполнять влагопотери. Наличие этих второстепенных с ландшафтных позиций
включений не значимо для птиц и мелких млекопитающих, но важно для земноводных,
- группы экологически более однообразной и на данной территории бедной видами.
Поэтому население первых двух классов лучше аппроксимируется более общими
ландшафтными (пейзажными) или геоботаническими параметрами среды, в то время
как информация о значимых для земноводных, но частных для Западно-Сибирской равнины,
особенностях местообитаний, отсутствует и не может быть получена по средне- и
мелкомасштабным географическим картам и космическим снимкам. Птицы и мелкие
млекопитающие, кроме того, занимают все ландшафтные урочища, в то время, как
земноводные не встречены в учетах на большей части тундровой зоны и в некоторых
местообитаниях лесостепи и степи. В связи с этим возникает неразрешимая
математически неопределенность в оценке сходства «нулевого» населения. Дело в
том, что коэффициент Жаккара-Наумова не учитывает сходства по отсутствующим
видам, как, например, коэффициент корреляции. Поэтому нулевые варианты имеют
нулевое сходство и соответственно сильное влияние зональных отличий в теплообеспеченности,
приводящее к отсутствию земноводных в тундрах не может быть оценено. Если его
принять за 100% (абсолютное сходство по отсутствию), подобное противоречие
возникает там, где численность мало отличима от нуля. В действительности сила и
общность связи факторов среды с неоднородностью населения земноводных значительно
выше приводимых значений, однако они не могут быть правильно оценены по чисто
математическим причинам.
Классификационные
представления и оценки связи природно-антропогенных режимов с неоднородностью
сообществ, наиболее информативны для населения птиц. Для мелких млекопитающих и
особенно для земноводных полнота объяснения существенно меньше.
Множественная
оценка связи всех отдельно взятых факторов и режимов значительно меньше суммы
их влияния, что связано с взаимной корреляцией отдельных факторов и их неразложимых
сочетаний. Специальное выявление относительно более слабых по силе влияния
факторов и режимов, позволило значительно увеличить полноту объяснения
территориальной изменчивости орнитокомплексов (Равкин, 1984). На основе
подобранной системы значимых структурообразующих факторов среды и
природно-антропогенных режимов с помощью линейной качественной аппроксимации и
обучающей матрицы коэффициентов сходства населения птиц была спрогнозирована
его неоднородность на необследованных территориях, по известным для них
факторам среды. При этом учтенная дисперсия расчетных показателей сходства при
интерполяции уменьшалась на 4-7% по отношению к объяснению неоднородности
обучающей выборки. Это свидетельствует о значительной надежности объяснения.
ЗАКЛЮЧЕНИЕ
Сопоставление
выявленных структур и классификаций можно проводить по набору и значимости
структурообразующих факторов, основным трендам и инвариантным блокам. Это
позволяет обобщать частные и формировать более общие представления о пространственной
неоднородности животного населения. Результатом анализа структурных
представлений могут быть принципы и правила формирования животного населения.
Они, с одной стороны, представляют собой способ организации материала, который
способствует анализу структур, формированию обобщенных представлений и
иллюстративности материала. С другой
стороны, эти построения характеризуют причины и последовательность
территориальных изменений животного населения. Опубликованные нами ранее
принципы и правила сформулированы при обобщенном описании сообществ птиц,
мелких млекопитающих и земноводных Западно-Сибирской равнины и Алтае-Саянской
горной страны (Равкин и др., 1994б). Некоторые из этих принципов и правил
сформулированы ранее другими исследователями вне связи с пространственно-типологической
структурой животного населения. Нами лишь прослежена их роль в формировании
населения. Эти, известные ранее принципы, перефразированы применительно к организации
сообществ. Пользуясь сформулированными принципами, можно отметить, что
пространственно-типологическая изменчивость облика населения птиц связана в основном с независимым замещением одних
видов другими и, в меньшей степени, - с обеднением-обогащением населения. У
мелких млекопитающих формирование сообществ идет по принципу обеднения-обогащения
и меньше определяется зависимым и независимым замещением видов, а также сменой
стаций. У земноводных доминирует обеднение-обогащение сообществ. Остальные
принципы и правила организации свойственны всем упомянутым группам животных.
Повторяемость и влияние типологического сходства четче всего прослеживаются в населении
птиц. Сообщества мелких млекопитающих испытывают большее влияние
хорологического сходства.
Таким
образом, пространственно-типологическая структура и организация сообществ на
одной и той же территории не одинакова для различных таксономических групп
животных, т.к. зависят не только от изменчивости значимых в данной местности
факторов, но и от специфики экологических требований видов.
Исследования,
послужившие основой для настоящего сообщения, поддержаны Российским фондом
фундаментальных исследований.
СПИСОК
ЛИТЕРАТУРЫ
Блинова Т.К.,
Блинов В.Н. 1997 и 1999. Птицы Южного Зауралья. Т. 1 и 2. Новосибирск: Наука.
296 и 288 с.
Вартапетов Л.Г. 1984. Птицы таежных междуречий. Новосибирск:
Наука. 242 с.
Вартапетов Л.Г.
1998. Птицы северной тайги Западно-Сибирской равнины. Новосибирск: Наука. 327
с.
Гынгазов А.М.,
Миловидов С.П. 1977. Орнитофауна Западно-Сибирской равнины. Томск: Изд-во
Томского ун-та. 351 с.
Равкин Е.С., Бышнев
И.И., Кочанов С.К. и др. 2001а. Пространственное разнообразие летнего населения
плиц Восточно-Европейской и Западно-Сибирской равнины // Достижения и проблемы
орнитологии Северной Евразии на рубеже веков. Казань. С. 214-238.
Равкин Е.С., Равкин
Ю.С., Вартапетов Л.Г. и др. 2001б. Классификация летнего населения птиц равнин
Северной Евразии // Сибирский экологический журнал. № 6. С.
Равкин Ю.С. 1978. Птицы лесной зоны Приобья. Новосибирск:
Наука. 288 с.
Равкин Ю.С. 1984.
Пространственная организация населения птиц лесной зоны. Новосибирск: Наука.
264 с.
Равкин Ю.С. 1991. Реализация и развитие зоогеографических
представлений А.П.Кузякина в Западной Сибири // Систематика животных,
практическая зоология и ландшафтная зоогеография. М.: Издательство Московского
общества испытателей природы. С. 47-58.
Равкин Ю.С. 1997. Особенности морфологического изучения
животного населения на зональных трансектах (исходные концепции и допущения) //
Известия АН. Серия биологическая. № 5. С. 603-612.
Равкин Ю.С., Богомолова И.Н., Ердаков Л.Н. и др. 1996.
Особенности распределения мелких млекопитающих Западно-Сибирской равнины /
Сибирский экологический журнал. Т. 3. № 3-4. С. 307-317.
Равкин Ю.С.,
Богомолова И.Н., Ердаков Л.Н. и др. 1997. Пространственно-типологическая
структура и организация населения мелких млекопитающих Западно-Сибирской
равнины // Успехи современной биологии. Т. 117. Вып. 6. С. 738-755.
Равкин Ю.С., Вартапетов Л.Г., Колосова Е.Н. и др. 1994а.
Видовое разнообразие птиц Западно-Сибирской равнины и особенности их летнего
распределения // Сибирский экологический журнал. Т. 1. № 6. С. 521-535.
Равкин Ю.С., Вартапетов Л.Г., Юдкин В.А. и др. 1994б.
Пространственно-типологическая структура и организация летнего населения птиц
Западно-Сибирской равнины // Сибирский экологический журнал. Т. 1. № 4. С.
303-320.
Равкин Ю.С., Вартапетов Л.Г., Юдкин В.А. и др. 1995.
Территориальная неоднородность населения земноводных Западно-Сибирской равнины
// Сибирский экологический журнал. Т. 2. № 2. С. 110-124.
Равкин Ю.С.,
Вартапетов Л.Г., Юдкин В.А. и др. 2001в. Классификация летнего населения птиц
Западной Сибири // Известия АН. Серия биологическая. № 3. С. 362-371.
Равкин Ю.С., Гуреев
С.П., Покровская И.В. и др. 1985. Пространственно-временная динамика животного
населения. Новосибирск: Наука. 207 с.
Равкин Ю.С.,
Лукьянова И.В. 1976. География позвоночных южной тайги Западной Сибири.
Новосибирск: Наука. 360 с.
Равкин Ю.С., Юдкин В.А., Вартапетов Л.Г. и др. 2000а.
Классификация населения птиц Западно-Сибирской равнины (вторая половина лета)
// Сибирский экологический журнал. Т. 7. Вып. 6. С. 743-754.
Равкин Ю.С., Юдкин В.А., Жуков В.С. и др. 2000б.
Классификация птиц Западно-Сибирской равнины по сходству распределения // Сибирский
экологический журнал. Т. 7. Вып. 3. С. 337-345.
Растительность Западно-Сибирской равнины (карта). 1976. ГУГК
СССР.
Терентьев П.В.
1959. Метод корреляционных плеяд // Вестник Ленингр. ун-та, серия биол. №. 9.
С. 137-141.
Философская энциклопедия, 1975. М. Т. 5. 740 с.
Ходашова К.С. 1966. О географических особенностях структуры
населения наземных позвоночных животных // Зональные особенности населения
наземных позвоночных животных. М. С. 7-38.
Чернов Ю.И. 1975. Природная зональность и животный мир суши.
М.: Мысль. 222 с.
Чернов Ю.И., Пенев Л.Д. 1993. Биологическое разнообразие и
климат / Успехи совр. биол. Т. 113. Вып. 3. С. 515-531.
Чернов Ю.И.,
Ходашова К.С., Злотин Р.И. 1967. Наземная зоомасса и некоторые закономерности
ее зонального распределения // Журн.общ.биол. Т. 28. № 2. С. 188-187.
Чупахин В.М. 1987. Основы ландшафтоведения. М.:
Агропромиздат. 183 с.
Jarvinen O., Sammalisto L.
1976. Regional trends in the avifauna of
Finish peatland bogs // Ann. Zool. Finnici. № 13. P.31-43.
Jarvinen O., Hayrinen U., Kouki
J. 1987. Reversed lalitudinal gradients in total density and species richness
of birds breeding on Finnish mires // Ornis fenn. 64. № 2. P. 67-73.
Kerr
J.T., Packer L. 1997. Habitat heterogeneity as a determinant of mammal species
richness in high-energy regions // Nature. Vol. 385/16. P. 252-254.
Ravkin Yu.S., Vartapetov L.G.,
Yudkin V.A. et al. 1995. Spatial structure of the Amphibian Assemblages of the
West Sibirian Plain / Amphibian Populations in the Commonwealth of Independent
Status: Current Status and Declines. - Moscow: Pensoft. P. 74-87.
РЕЗЮМЕ
ПРОСТРАНСТВЕННО-ТИПОЛОГИЧЕСКАЯ
ОРГАНИЗАЦИЯ ЖИВОТНОГО НАСЕЛЕНИЯ ЗАПАДНО-СИБИРСКОЙ РАВНИНЫ
(НА ПРИМЕРЕ ПТИЦ,
МЕЛКИХ МЛЕКОПИТАЮЩИХ И ЗЕМНОВОДНЫХ)
Ю.С.Равкин
Общий
характер выбора местообитаний у птиц и мелких млекопитающих на Западно-Сибирской
равнине определяется, в первую очередь, зональностью и застроенностью; у птиц,
кроме того, - наличием озёр и рек. Обилие земноводных тоже зависит от зональности,
но не менее значима степень увлажнения и, также как у мелких млекопитающих, влияние половодий.
Общим значимым фактором среды для всех трех групп животных является
продуктивность биоценозов. Тенденция снижения плотности животного населения к
северу от степей в значительной степени нарушается за счет миграций птиц и
степенью распашки территории в южной части равнины, которая отрицательно влияет
на все три рассматриваемые группы животных, а также, возможно, за счёт
конкурентных отношений в населении земноводных. Видовое богатство населения
наиболее высоко в лесостепи и почти неуклонно убывает к северу и югу с
единственным отклонением в подтаежных лесах, где число отмеченных видов меньше,
чем в южной тайге. Это связано с небольшой шириной подтаёжной подзоны и
невысоким разнообразием образующих ее местообитаний. Для всех трёх групп
животных наиболее значима связь с зональностью (подзональностью), несколько
меньше - с облесенностью и составом лесообразующих пород. Сила и общность связи
территориальной изменчивости сообществ с остальными факторами среды, в том
числе и с антропогенным воздействием, существенно меньше. Последнее имеет
высокое локальное влияние, но в целом по Западно-Сибирской равнине мало
значимо. Пространственно-типологическая изменчивость облика населения птиц
связана в основном с независимым
замещением одних видов другими и, в меньшей степени, - с обеднением-обогащением
населения. У мелких млекопитающих формирование сообществ идет по принципу
обеднения-обогащения и меньше определяется зависимым и независимым замещением
видов, а также сменой стаций. У земноводных доминирует обеднение-обогащение сообществ.
Повторяемость и влияние типологического сходства четче всего прослеживаются в
населении птиц. Сообщества мелких млекопитающих испытывают большее влияние хорологического сходства.
SPACE-TYPOLOGICAL ORGANIZATION OF THE
ANIMAL COMMUNITY OF THE WESTERN SIBERIA PLAIN (BIRDS, SMALL MAMMALS AND
AMPHIBIANS TAKEN AS AN EXAMPLE)
Yu.S. Ravkin
The habitat generally chosen by birds and small
mammals of the Western Siberia Plain is primarilly determined by zonation and
urbanization degree, and additionally by the presence of lakes and rivers in
case with birds. The abundance of amphibians also depends on zonation, however
the moistening degree and (like in small mammals) flood influence are of no
less importance. The important environmental factor for the three groups of
animals is a biocenosis productivity. The tendency of decreasing of animal
community density north of steppes is considerably broken due to birds
migration and plowing areas in the southern part of the plain, that negatively
influence the three animal groups discussed, as well as probably due to the
competition within the amphibian community. The variety of community species is
richest in forest steppe, steadily decreasing to the north and south, with a
single deviation in subtaiga, where the number of marked species is less than
in south taiga. This is due to a small width of the subtaiga subzone and a low
diversity of its habitats. For all three groups of animals, most important is
the association with zonation (subzonation), somewhat less essential is that
with forest areas and forest-forming species composition. The association
between community area variability and other environmental factors, including
anthropogenic impact, is considerably less strong and common. The latter has a
great local impact, though it is not very important over the Western Siberia
Plain, as a whole. The space-typological variability of a bird community is
mainly associated with the independent replacement of some species and, to a
lesser degree, with impoverishment/enrichment of the community. In small
mammals, a community is formed according to the impoverishment/enrichment
principle, being less determined by a dependent and independent replacement of
species, and station alternation. In amphibians, an impoverishment/enrichment
of communities prevails. The repeatability and impact of a typological
similarity is most clearly observed in bird communities. Small mammals
communities are characterized by a greater influence of a chorologic
similarity.